
Remember me

Asthma, atopic dermatitis (AD), Chronic urticaria (CU), and food allergies are among the most common immunologic disorders managed by allergists. Asthma is a respiratory disease of the lower airway characterized by variable symptoms such as cough, wheezing, shortness of breath and bronchial hyperreactivity. Several asthma endotypes have been described including Th2-high asthma, characterized by high levels of type 2 cytokines and eosinophilic airway inflammation and Th2-low (or non-Th2) asthma associated with neutrophilic or paucigranulocytic inflammation [1]. AD is a widespread, chronic inflammatory skin condition marked by itchy, red, and dry skin. The causes of AD are multifactorial including environmental triggers, genetic predisposition, a compromised skin barrier and immune system dysregulation [2]. As with asthma, distinct pathophysiological mechanisms exist for AD, however, defining of reproducible endotypes remain challenging [3]. Chronic urticaria, a mast cell-driven inflammatory skin disorder, is characterized by recurrent wheals, angioedema, or both, lasting for more than six weeks [4]. CU can be classified as chronic spontaneous urticaria (CSU) and chronic inducible urticaria (CIndU) with CSU occurring in the absence of identifiable physical or environmental stimuli. In recent years, advances in immunological research have revealed that autoimmunity plays a central role in the pathogenesis of CSU [5]. Food allergies are abnormal immune responses triggered by exposure to specific foods and are classically defined as immunoglobulin E (IgE)-mediated hypersensitivity reactions. However, recent advances, e.g. in diagnostic tools, have led to the proposal of distinct food allergy endotypes [6].
Affecting 30–40% of the global population [7], these conditions are becoming increasingly prevalent and complex, often involving multifactorial triggers and overlapping phenotypes. While traditionally viewed as disorders of hypersensitivity to exogenous antigens (allergy), emerging research has revealed that some diseases may also involve immune responses directed against self-antigens, a phenomenon termed autoallergy. This concept, situated at the intersection of allergy and autoimmunity, remains underrecognized in daily clinical practice. In this review, we aim to provide allergists with a concise and clinically relevant overview of autoallergy by addressing five key questions (Table 1): (1) What is autoallergy and how does it differ from classical allergy? (2) How common is autoallergy? (3) Is autoallergy clinically relevant? (4) How can autoallergy be diagnosed? and (5) How is autoallergy treated? In this review, we focus on asthma, AD, CU, and food allergy, while excluding conditions like systemic lupus erythematosus and bullous pemphigoid, where IgE autoantibodies have been described but are less central to allergy practice and have been discussed in a prior publication [8].
Table 1 Summary: Key questions and answersWhat is Autoallergy and how Does it Differ from Classical Allergy?IgE is best known as a key effector in type I hypersensitivity reactions [9]. Classic IgE-mediated allergy involves sensitization to environmental (exogenous) antigens (e.g. pollen and food proteins), where antigen-presenting cells present these allergens to naive T cells. Differentiated Th2 cells activate B cells, initiating class-switch recombination and subsequent IgE production. IgE can bind to effector cells like basophils and mast cells via its high affinity receptor (FcεRI). Upon re-exposure, the allergen crosslinks surface-bound IgE on mast cells and basophils, triggering degranulation and release of histamine, leukotrienes, and cytokines [10].
Accumulating evidence has revealed that IgE, in addition to exogenous antigens, can also target self-antigens leading to autoimmune responses known as autoallergy, or type I autoimmunity [5]. In autoallergy, the immune system produces IgE autoantibodies directed against endogenous antigens also called autoantigens or autoallergens, including thyroid peroxidase (TPO), interleukin-24 (IL-24), manganese superoxide dismutase (MnSOD), among others [8]. As in classical allergy, this form of autoimmunity results in activation of mast cells and basophils through crosslinking of FcεRI-bound IgE, leading to inflammation and clinical manifestations such as wheals [5].
Endogenous IgE responses may arise through several immunological mechanisms that are increasingly recognized in chronic inflammatory and autoimmune conditions. One such mechanism is molecular mimicry, in which exogenous antigens (such as microbial or parasitic proteins) share structural or sequential similarities with tissue-specific human proteins or external allergens, leading to cross-reactive IgE responses11,12,13,14. For instance, Sánchez et al. proposed that TPO may resemble endogenous eosinophil proteins like eosinophil peroxidase, potentially triggering autoreactive IgE responses [12,13,14]. IgE reactivity to human profilin, the first IgE autoantigen to be identified, has been found in individuals with birch pollen allergy, suggesting molecular mimicry between plant and human homologs [15].
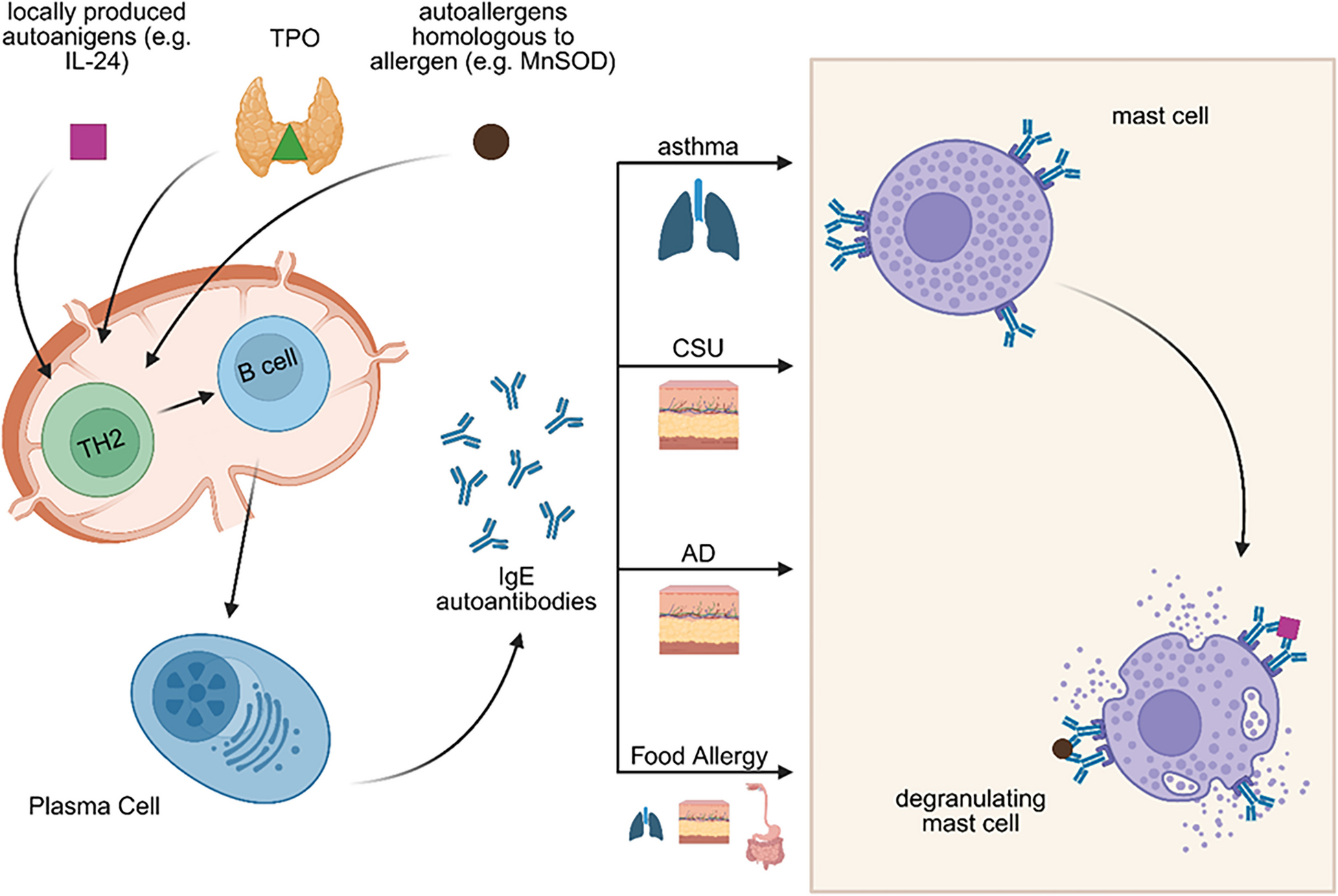
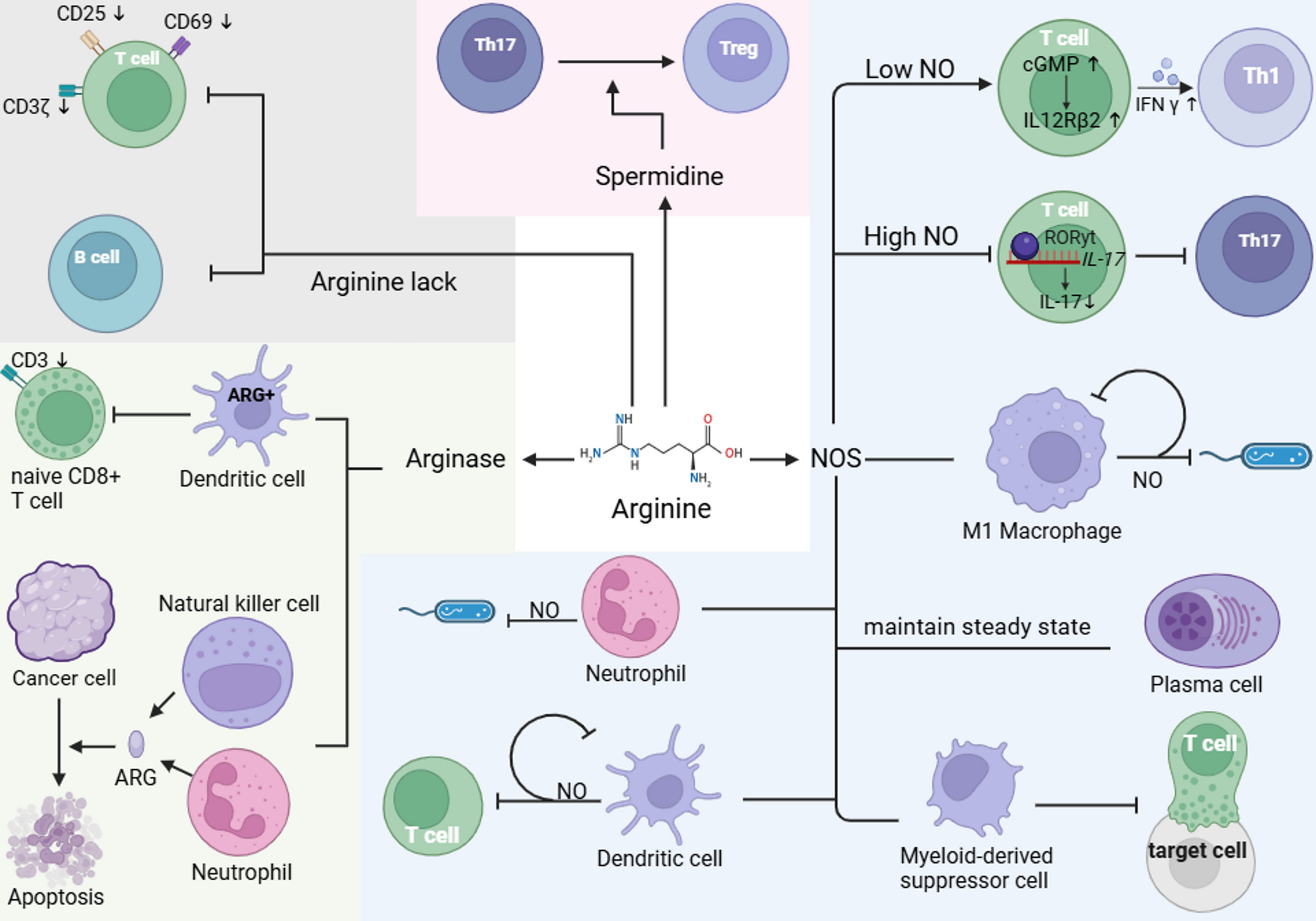
Another contributing mechanism is epitope spreading, where an initial immune response against a specific epitope gradually extends to involve other epitopes on the same or related self-antigens, further amplifying autoimmunity and autoallergy [12,13,14]. Additionally, local class-switch recombination to IgE may occur within tertiary lymphoid structures that develop in chronically inflamed tissues such as the skin. These ectopic lymphoid formations create a specialized microenvironment that supports ongoing B cell activation and class switching in situ, enabling the production of IgE against local tissue antigens independently of central immune organs [16, 17]. Such local IgE production has also been proposed in barrier organs, such as the respiratory epithelium and nasal mucosa, particularly in asthma and allergic rhinitis [18,19,20,21]. A proposed model of autoallergy (Fig. 1) mirrors the classical type I hypersensitivity cascade, with a sensitization phase followed by an effector phase, but with key differences in antigen origin.
Fig. 1
Pathogenesis of autoallergy in asthma, CSU, AD and Food Allergy. Similar to type I hypersensitivity reactions, autoallergy is also characterized by a two-step process consisting of a sensitization phase followed by an effector phase. However, the precise mechanisms underlying sensitization in autoallergy remain unknown. Several hypotheses have been proposed to explain the development of autoreactive IgE. One suggests that in the presence of alarmins, self-antigens may be taken up by antigen-presenting cells, leading to the activation of Th2 cells. These Th2 cells then promote the production of autoreactive IgE antibodies by plasma cells. Another proposed mechanism is molecular mimicry, in which structural similarities between environmental or microbial antigens and self-proteins lead to the production of IgE autoantibodies. IgE autoantibodies can bind to tissue-resident mast cells: in the lungs in asthma, in the skin in atopic dermatitis (AD), and in the lungs, intestines, and skin in food allergy. Mast cell activation and subsequent degranulation may be triggered by autoallergens that are cross-reactive with exoallergens (e.g. MnSOD), by re-exposure to locally produced autoallergens (e.g. IL-24), or by local allergens that cross-react with other human proteins (e.g. TPO and EXP). Created with BioRender.com
Functional assays, such as the basophil activation test (BAT), and in vivo tests, e.g. skin prick tests and passive transfer experiments, underscore the contribution of autoreactive IgE to pathogenesis of asthma, AD, CSU, and food allergy. Notably, passive transfer of IgE against thyroid peroxidase (TPO) from CSU patients has been shown to induce wheal-and-flare reactions in healthy skin, as demonstrated in the study by Sanchez et al., further supporting the pathogenic role of autoreactive IgE in CSU [22]. Basophils from IgE anti-eosinophil cationic protein (ECP) and IgE anti-eosinophil peroxidase (EXP) positive asthmatic patients, for instance, showed a dose dependent activation after stimulation with the culprit autoantigen [13]. Similar, assays using rat basophilic leukemia cells demonstrated that human α-lactalbumin triggers mediator release in sensitized milk allergic individuals [23].
How Common is Autoallergy?Autoallergy has been detected in a range of immune-mediated disorders, yet the prevalence of autoreactive IgE varies substantially depending on the underlying disease, the specific autoallergen, the characteristics of the study population, and the detection methods employed. Table 2 provides an overview of IgE autoreactivity across the reported disease entities, including information on identified autoantigens, prevalence, and detection techniques.
Table 2 Prevalence of IgE autoantibodies in atopic diseasesStudies on asthma have identified IgE autoantibodies targeting various autoantigens, including DFS70 [24], periplakin [25], rheumatoid factor [26], platelet antigens [27], EXP and ECP [13] with reported prevalence rates ranging from 8.7% to 70%. In the context of food allergy, existing reports on IgE autoantibodies are limited to cow’s milk allergy, where autoreactive IgE has been detected against human homologs of α-lactalbumin, β-casein, and κ-casein (see Table 2). Notably, the prevalence of autoreactive IgE in milk allergy appears to be high, ranging from 35% to 100% [23, 28,29,30].
The concept of autoimmunity in AD has been recognized for several decades, with studies dating back over eighty years identifying autoreactivity to human dander in patients with eczema [31, 32]. Thereafter, multiple studies [24, 33,34,35,36,37,38,39,40,41] have confirmed the presence of autoreactive IgE in individuals with AD, with at least 140 [11] different self-antigens identified. Examples of human autoantigens in AD include autologous sweat, Homo sapiens allergen 1–5 [42, 43], and MnSOD [44]. Prevalence studies on autoreactive IgE in AD patients have shown a broad range, from 23% to as high as 91% [8], depending on the sample size and methodology. Larger-scale research typically reported prevalence rates around 20–30% [8]. Of note, a recent analysis found autoreactive IgE in 16.4% of those with both AD and other atopic conditions and only 9.6% in those with only AD [45].
In CSU, several IgE autoantibody targets have been identified, including IL-24, TPO, thyroglobulin (TG), double-stranded DNA (dsDNA), ECP, EXP, FcεRI, tissue factor (TF) and tissue-transglutaminase 2 [46,47,48,49, 96]. Reported prevalence rates of autoreactive IgE in CSU vary considerably and range from 0–100% [8]. However, the majority of studies report a prevalence between 30–60% [8, 48]. A recent study involving 111 CSU patients showed that more than half of the patients (58%) exhibited an autoallergic phenotype, suggesting that autoallergy is a common feature of CSU [50].
Autoreactive IgE can also be detected in some healthy individuals, with reported prevalence rates ranging from 0 to ~30% (see Table 2). However, in these cases, such antibodies are generally regarded as non-pathogenic. For instance, Sánchez et al. demonstrated that healthy subjects with detectable IgE against ECP and EXP showed minimal to no basophil activation upon stimulation with these autoantigens [13, 14].
Is Autoallergy Clinically Relevant?The clinical significance of autoallergy is based on the evidence linking autoreactive IgE with disease severity and other clinical features. In AD, IgE autoantibodies have been also linked to dry skin, dyshidrosis, itchyosis, infections and pruritus [38]. In asthma, patients with IgE anti-periplakin had a higher frequency of nasal polyps than patients without autoantibodies [25]. In CSU, IgE anti-TPO has been associated with higher frequency of atopy and asthma [51]. Additionally, IgE anti-TPO levels correlated with IgG anti-TPO levels [52, 53] supporting the notion that CSU patients are at risk for autoimmune thyroid diseases and thyroid dysfunction [5]. Cross-reactivity between bovine and human proteins is particularly clinically relevant when infants with cow’s milk allergy remain symptomatic despite maternal avoidance of dairy [23, 54, 55].
In CSU, IgE autoantibody levels were associated with disease activity in some studies [50, 56], but not in others [22, 52, 53]. For example, IgE anti-IL-24 levels correlated with the urticaria activity score over 7 days (UAS7) [56]. Further, it has been shown that autoantibodies are elevated during exacerbation periods [51]. Also, in AD autoreactive IgE correlates with disease severity [37, 38, 41, 44], while some studies failed to find this correlations [
Comments (0)