
Remember me


You have full access to this article via your institution.
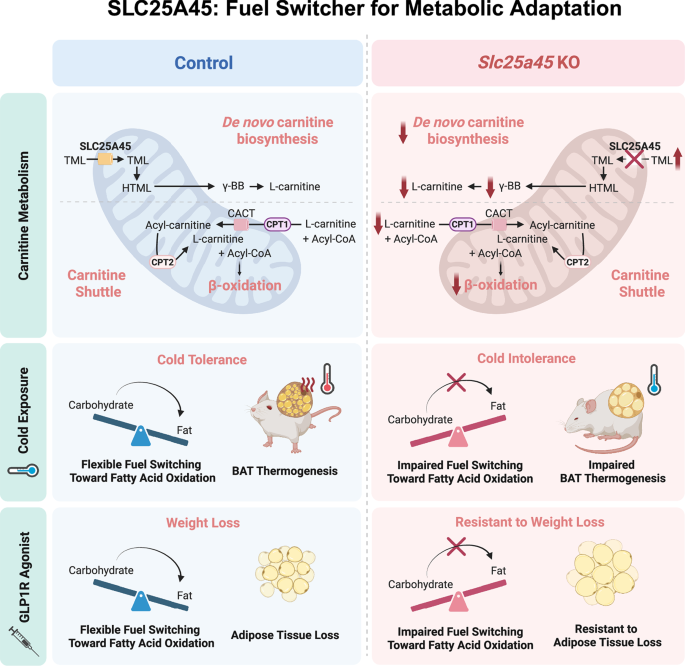
Nuclear cGAS acts as a vital line of defense against DNA virus infections. In a recent Cell paper, Lahaye and colleagues uncover a novel mechanism in which nuclear cGAS detects viral infection, triggered by the amplification of centromeric DNA.
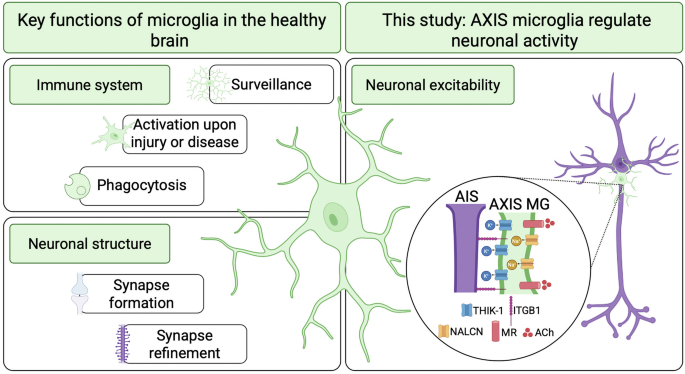
Understanding antiviral immunity is crucial for developing effective treatments and overcoming viral resistance. One key component of this response is the DNA sensor cyclic GMP-AMP synthase (cGAS) which detects the abnormal presence of DNA in the cytoplasm and produces cyclic GMP-AMP (cGAMP). cGAMP then activates stimulator of interferon genes (STING), initiating downstream signaling that results in the production of type I interferons and pro-inflammatory cytokines, communication with bystander cells, and ultimately, cell death. This pathway enables the detection of foreign pathogens and triggers the innate immune response. However, most DNA viruses enter the nucleus as part of its infection cycle. Fortunately, cGAS is also present in the nucleus,1 but remains in an inert form, often through tight association with the nucleosome core to prevent self-DNA recognition and unintended immune activation2,3,4 (Fig. 1a). How nuclear cGAS turns active from an inert state upon virus invasion is a subject of intensive study. Studies in human immunodeficiency virus (HIV) and herpes simplex virus 1 (HSV-1) suggest that nuclear cGAS directly detects viral DNA.5,6 Now, Lahaye et al.7 proposes a novel mechanism in which nuclear cGAS is activated by self-host DNA upon viral infection.
Fig. 1: Host antiviral defense via TLS-mediated activation of nuclear cGAS at centromeres.
a In uninfected cells, nuclear cGAS is enriched at centromeres but remains inactive due to its tight association with nucleosomes. b During herpesvirus infection, the viral E3 ligase ICP0 accumulates at centromeres and promotes TLS-mediated amplification of centromeric DNA. This process requires ICP0’s ubiquitin ligase activity and the host deubiquitinase USP7, which may counteract ICP0 auto-ubiquitination. In contrast, the viral deubiquitinase UL36USP inhibits TLS-mediated centromeric DNA amplification. The resulting increase in centromeric DNA activates cGAS, triggering cGAMP synthesis and a downstream interferon response.
Previous studies have found that nuclear cGAS is enriched at the centromere,8 a vital part of our genome important for chromosome segregation during cell division. Interestingly, HSV-1 also associates with the centromere during viral infection.9 Furthermore, HSV-1 encodes viral protein infection cell protein 0 (ICP0), which induces proteasomal degradation of centromere proteins, leading to destabilization of centromere architecture.10 Lahaye and colleagues investigate the possibility that ICP0 activates nuclear cGAS at the centromere. Experimentally, the authors expressed ICP0 in human blood monocytes and assessed its effect following their differentiation into dendritic cells, a known target of HSV-1 infection. They found that expression of ICP0, but not its mutant deficient for ubiquitin ligase activity (ICP0 FXE), activated cGAS signaling and production of interferons. They further showed that depletion of cGAS abolished the production of interferons in ICP0-expressing cells, confirming that cGAS signaling was indeed activated.
Since nuclear cGAS can be found at the centromere, this raises an intriguing possibility that the activation of cGAS is triggered by host centromere DNA during HSV-1 infection. The authors found that the centromere DNA was more accessible in cells expressing ICP0, as demonstrated by multiple methods including genome-wide sequencing, quantitative PCR and fluorescence microscopy. Further whole-genome sequencing revealed amplification of centromere DNA in cells expressing ICP0, but not mutant ICP0 FXE. Interestingly, this amplification of DNA only occurred at the centromere, and no amplification of DNA from mitochondria and from chromosome arms was observed.
In normally cycling cells, DNA is amplified during the S phase through replication. Typically, replication of centromere DNA is aided by translesion DNA synthesis (TLS).11 Consistent with this, the authors found that TLS is also required for the amplification of centromere DNA in ICP0-expressing cells, despite their non-cycling state. Knocking out key components of the TLS machinery, PCNA, POLH, REV3L and POLD1, was sufficient to inhibit centromeric DNA amplification and cGAS activation in these cells. Ubiquitin-specific peptidase 7 (USP7), a known TLS regulator and a host target of ICP0, was also shown to play a critical role.
Notably, virus-induced centromeric DNA amplification and associated cGAS activation is not limited to HSV-1. The authors demonstrated that multiple functional homologs of ICP0 from different herpesviruses, including ORF63 in equine herpesvirus, BICP0 in bovine herpesvirus, and IE1 from human cytomegalovirus, all elicit a similar response to ICP0. Thus, paradoxically from a virus’s standpoint, ICP0 and its counterparts promote a host antiviral defense mechanism.
Lahaye et al., however, also found that HSV-1 can evade the cGAS antiviral response by preventing TLS. Mechanistically, the authors show that HSV-1 encodes a small deubiquitinase UL36USP which inhibits TLS and, in doing so, blocks centromeric DNA amplification and subsequent activation of cGAS signaling.
In summary, Lahaye and colleagues have revealed a novel host defense system against viral infection based on viral-induced centromere amplification and recognition (VICAR) (Fig. 1b). The authors provide the first evidence that the centromere is involved in innate immunity and acts as a detection tool against viral proteins. Viral proteins can promote TLS by ubiquitination, amplifying centromere DNA, which then triggers cGAS activation. This leads to downstream production of interferons and starts an innate immunity response.
An expansive model is proposed through the experimental data. However, several questions remain unresolved. For instance, the molecular mechanisms by which ubiquitination events, involving the viral proteins ICP0 and UL36USP, along with the host protein USP7, regulate TLS are still unclear. It is also unknown whether similar mechanisms operate across different cell types upon HSV-1 infection. Another intriguing question is whether events associated with the centromere amplification provide any advantage to DNA virus during latency or reactivation. Conversely, could they also contribute to the rapid evolution of centromeres in host cells, for example through viral genome integration? Finally, the findings raise a broader question of whether centromere amplification and cGAS activation extend to other pathological conditions, especially those involving non-dividing, post-mitotic cells.
In the landscape of viral–host cell interaction, the centromere appears to be a molecular minefield. One viral protein (ICF0) can trigger the “mine” (cGAS) embedded at centromeres, while another (UL36USP) disarms it by suppressing this response. This interplay highlights an exciting therapeutic opportunity: for example, targeting UL36USP could evoke the host’s innate immune response against related DNA viruses. Future development of antiviral strategies that exploit VICAR may pave the way for novel therapeutic treatment.
Comments (0)