
Remember me
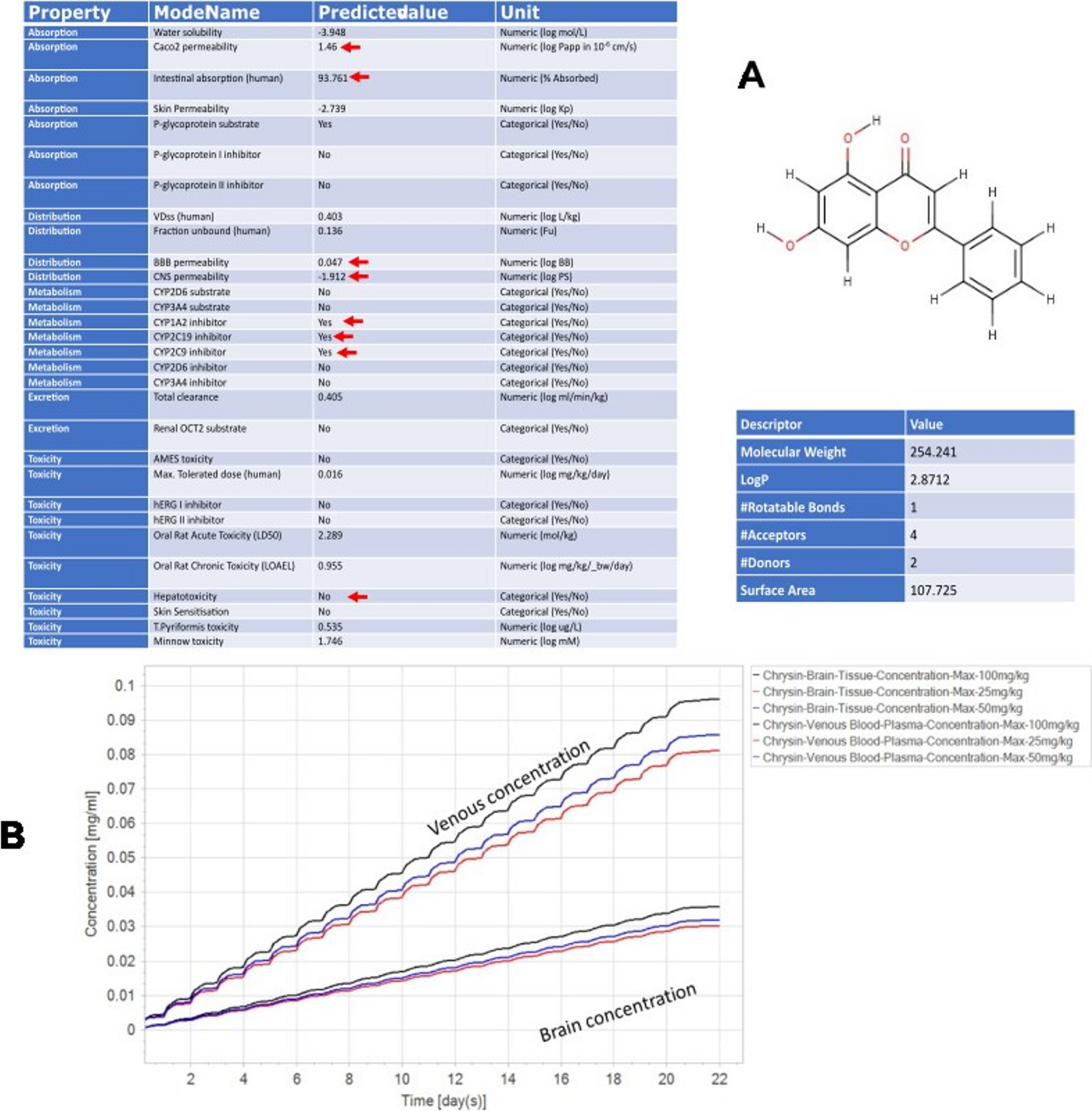

Neurogenesis is the process of generating new neurons through an orchestrated series of transcriptional and morphological modifications from neural progenitor cells (NPCs) (Bond et al. 2015; Silva Siqueira et al. 2021). Neurogenesis occurs from the development until adult life, including several phases: proliferation, migration, differentiation, and circuit integration of mature neurons (LiCausi and Hartman 2018; Araki et al. 2021). Moreover, Klotho-induced neurogenesis in the postnatal phase enhances neuroplasticity and hippocampal-dependent cognition in mice and rats (Dubal et al. 2014; Salech et al. 2019; Laszczyk et al. 2017; Mytych 2022). Furthermore, dysregulation of embryonic neurogenesis can lead to malformations and functional disturbances characteristic of neurodevelopmental disorders. Therefore, the proper progression of embryonic neurogenesis supports healthy neurodevelopment (Villalba et al. 2021). In this context, a recent study has demonstrated that Klotho promotes embryonic neurogenesis. Moreover, Klotho-induced neurogenesis in the postnatal phase enhances neuroplasticity and hippocampal-dependent cognition in mice and rats (Dubal et al. 2014; Salech et al. 2019; Laszczyk et al. 2017; Mytych 2022). Therefore, Neurogenesis plays an important role in CNS homeostasis, highlighting its potential as a therapeutic target for neurodegenerative and neuropsychiatric disorders (Prokhorova et al. 2019; Vo et al. 2018). Table 1 summarizes the main findings about the role of Klotho in neurogenesis in the literature.
Table 1 Summary of the main results found in studies of klotho’s role in neurogenesisKlotho and Embryonic Neurogenesis and OligodendrogenesisIn the developing brain, radial glia from the ventricular zone (VZ) divide into intermediate progenitors, which amplify and migrate out of the VZ to establish at the SVZ, where these progenitors divide and generate neuroblasts. Thus, the neuroblasts migrate through the cortex. In embryonic neurogenesis, neural stem cells (NSCs) divide to generate differentiated progenitor cells and maintain the stem pool (Engler et al. 2018).
In 2017, Laszczyk’s study group investigated the expression of Klotho during development using qPCR on the mRNA of mouse NPCs, detecting its first expression at 17 embryonic days (Laszczyk et al. 2017). Also, it was demonstrated that Klotho deficiency did not impair embryonic neurogenesis. In this study, only at 7 weeks postnatally, Klotho-deficient mice showed a decrease in the number of NPCs and immature neurons compared to wild-type mice. In addition, the study showed that hippocampal volume at 3 and 7 weeks of age was not significantly different between WT and KO mice. However, at 7-week-old KO mice exhibited a decrease in fimbria volume (Laszczyk et al. 2017). Furthermore, α-Klotho mutant mice show premature biochemical and morphological changes in synaptic proteins and terminals, microtubules, and organelles, as well as reduced brain and spinal cord volume. It is possible that this reduction in brain volume was caused by accelerated degeneration or developmental defects (Shiozaki et al. 2008).
Nevertheless, a recent study by Kim et al. demonstrated that α-Klotho may also play a role in embryonic neurogenesis and oligodendrogenesis. This study showed that α-Klotho is expressed in NPCs since the 13.5-day embryonic mouse. In addition, Klotho knockdown in embryonic NPCs reduced the proliferation rate, decreased the neurosphere diameter, and decreased the number of NPCs labeled with Ki67 (a proliferation marker). Furthermore, knockdown of α-Klotho repressed signaling pathways involving Akt and ERK phosphorylation, suggesting that this signaling may represent a possible mechanism by which Klotho promotes the proliferation of mouse embryonic NPCs (Kim et al. 2025). Moreover, knocking down α-Klotho also impaired the differentiation of NPCs into neurons or oligodendrocytes, but not astrocytes. In addition, a terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay showed that Klotho deletion increased basal cell apoptosis. This suggests that α-Klotho is an important factor in the survival of neural stem cells (Kim et al. 2025).
In contrast, overexpressing α-Klotho in transmembrane or secreted form in embryonic cells enhances the proliferation of NPCs (number of neurospheres and Ki67 labeling). Furthermore, Klotho overexpression also promotes neuron and oligodendrocyte differentiation, resulting in an increased number of neurons with highly dendritic arborization. In conclusion, Kim’s study indicates that α-Klotho promotes cell survival, NPC proliferation, and neuronal and oligodendrocyte differentiation (Kim et al. 2025). Therefore, Klotho also may play a modulatory role in neuronal fate determination from embryonic stem cells in neurogenesis and oligodendrogenesis.
Klotho and Adult Hippocampal NeurogenesisThe adult hippocampal neurogenesis (AHN) evolves the neurogenic niche, which is localized in the SVZ of the lateral ventricles and in the subgranular zone (SGZ) of the hippocampal dentate gyrus (DG). The niche comprises cellular and non-cellular components that influence the fate of the NPC: self-renewal or differentiation (Kempermann et al. 2004; Li Ming and Song 2011). In this process, a bipotent radial glia-like stem cell (type-1 cell) has undifferentiated molecular markers such as nestin and glial fibrillary acidic protein (GFAP). Type 1 divides symmetrically for long-term expansion. However, type-1 cells can divide asymmetrically to give rise to lineage-committed progenitor cells (type-2 and type-3 cells), which subsequently differentiate into immature neurons or astrocytes. Some type 2 cells and all type 3 cells express doublecortin (DCX), a marker for immature neurons, while intermediate progenitors amplifying express SOX2 + and Ki-67. Ultimately, immature neurons differentiate into mature neurons as they migrate to the granule cell layer and begin to express NeuN, as shown in Fig. 2 (Salech et al. 2019; Laszczyk et al. 2017; Bond et al. 2015; Kempermann et al. 2004; Li Ming and Song 2011). The SGZ contains neural stem cell pools that generate new neurons throughout postnatal life, which are crucial for maintaining structural integrity, regenerating brain tissue, and promoting neuroplasticity (Isaev et al. 2019). This process ensures controlled cell expansion and neuronal formation, and prevents depletion of the NPC pool (Bonafina et al. 2020; Su et al. 2019).
Fig. 2
Klotho modulates distinct phases of hippocampal neurogenesis. The figure illustrates the stages of neurogenesis: proliferation, neuronal differentiation, and subsequent neuronal maturation
In 2014, Cliton’s group investigated Klotho mRNA expression in postnatal developing and adult brains of rats with in situ hybridization. The analyses detected Klotho expression in many brain structures of gray matter as early as the first postnatal day. Additionally, it has been demonstrated that the KL gene is highly expressed in the DG, particularly in mature neurons (Clinton et al. 2013). Also, the literature has reported that Klotho overexpression may enhance hippocampus-dependent cognition by modulating adult neurogenesis, which probably influences the cellular decision of NPCs differentiation into neuroblasts through asymmetric division (Dubal et al. 2014; Salech et al. 2019; Laszczyk et al. 2017; Su et al. 2019).
Salech et al. showed that AHN is increased in KL knockout mice. In contrast, local downregulation of Klotho in the mouse hippocampus impairs the AHN process, leading to a decrease in hippocampus-dependent cognition. However, treatment with recombinant Klotho increases the proliferation rate of cultured NPCs and promotes their differentiation into neurons (Salech et al. 2019). The site-specific deletion of Klotho in the hippocampus generates distinct responses in the neurogenesis compared to whole-body deletion of the KL gene in mice. A possible hypothesis for this difference is that whole-body deletion may have generated compensatory responses that promote cell proliferation in this model, since second metabolites may cross the barrier. Furthermore, the hippocampus is a region composed of a neurogenic niche. Therefore, the local knockdown of Klotho may have a more direct influence on neurogenesis, impairing hippocampal-dependent memory (Salech et al. 2019).
Furthermore, posterior studies have reported that Klotho prevents spatial memory loss by promoting hippocampal neurogenesis in NPCs of postnatal mice (Dubal et al. 2014; Salech et al. 2019; Laszczyk et al. 2017; Vo et al. 2018). Laszczyk’s group has reported that KL gene deletion in hippocampal NPC decreased the number and size of neurospheres, as well as the number of immature neurons, delayed neuron maturation, and impaired neuronal dendritic arborization. Therefore, Klotho deficiency in mice impaired the proliferation process and compromised neuronal maturation, suggesting an early depletion of the neurogenic niche. In this context, supplementation with recombinant Klotho could reverse the decrease in NPCs proliferation, protect against premature aging of the niche, and mitigate spatial discrimination impairment. In contrast, Klotho overexpression increased the NPCs’ proliferation rate, the number of immature neurons, and enhanced neuronal dendritic arborization (Laszczyk et al. 2017; Vo et al. 2018). These results suggest that Klotho is essential to modulate different stages of adult neurogenesis.
Moreover, the results from Dubal’s group study in 2014 suggest that Klotho overexpression enhances the expression of the immature neuron glutamate receptor subunit, GluN2B. Elevated GluN2B protein levels were associated with neuroplasticity, since these receptors facilitate long-term potentiation (LTP) and improve cognitive ability in young mice. Therefore, since the N-Methyl-D-Aspartate (NMDA) receptor modulates the proliferation and differentiation of immature neurons, increasing GluN2B subunits could improve neurogenesis (Dubal et al. 2014).
Other studies have demonstrated an association between Klotho expression and the promotion of neurogenesis (Dias et al. 2021). Dias’ research group showed that female mice subjected to intermittent fasting exhibited an improvement in long-term memory retention, as well as a higher number of proliferating NPCs and neuroblasts, which were associated with an upregulation of the KL gene in the hippocampus (Dias et al. 2021). Furthermore, a similar association was explored in a model of caloric restriction and treatment with Astragalus, which is an antioxidant and anti-inflammatory herbal compound. In this study, increased neurogenic markers, such as doublecortin (DCX) and NeuN, and brain-derived neurotrophic factor (BDNF) levels, as well as improved cognitive performance, were associated with an enhancement of Klotho expression in the hippocampus (Setel et al. 2022). Similarly, a recent study has shown that treatment with recombinant soluble Klotho increases the number of immature neurons in a histological analysis of mice (Roig-Soriano et al. 2025). In contrast, Ho’s 2020 study shows decreased synaptic plasticity in the DG and CA1 regions of the hippocampus, as well as reduced neural progenitor cells in the DG, associated with decreased Klotho levels in C9orf72 knockout mice that simulate amyotrophic lateral sclerosis and frontotemporal dementia (Ho et al. 2020).
Since Klotho is a co-receptor of the FGF-23 receptor and performs part of its functions through receptor activation, FGF-23 deletion can also induce a hippocampal-dependent cognitive impairment in mice. However, FGF-23-deficient mice exhibited minor changes in hippocampal neurogenesis, characterized by decreased neuronal differentiation without significant differences in the proliferation phase (Laszczyk et al. 2019). These results suggest that FGF-23 deletion could not be related to neurogenesis impairment, whereas Klotho deficiency is associated with cognitive impairment due to an imbalance in neurogenesis. In conclusion, an increase in Klotho gene expression or circulating protein enhances pro-neurogenic parameters and their respective functional consequences. In contrast, lower levels of Klotho are associated with impaired adult neurogenesis.
In 2019, Su et al. demonstrated, using single-cell RNA sequencing, that α2-chimaerin (a Rho GTPase-activating protein) is essential for maintaining the NPC pool during adult neurogenesis. In this study, a subpopulation of NPCs with high Klotho expression in the hippocampus of α2-chimaerin knockout mice was shown to provide signals that modulate the proliferation rate of NPCs (Su et al. 2019). Moreover, decreased Klotho expression in the NPCs population is associated with premature cell differentiation and loss of the NPCs pool, which impairs synaptic plasticity and compromises hippocampal function (Dubal et al. 2014; Laszczyk et al. 2017; Vo et al. 2018; Su et al. 2019). These results corroborate previous studies demonstrating that Klotho deficiency decreases NPC proliferation and causes early depletion of the neurogenic niche associated with aging-related diseases in the CNS (Laszczyk et al. 2017; Vo et al. 2018; Su et al. 2019). Therefore, the data summarized in the diagram in Fig. 2 corroborate the hypothesis that the Klotho protein is important for cellular decision-making and the preservation of the NPCs pool during neurogenesis.
The scheme shows that higher levels of Klotho expression in KL-overexpression mutant mice, endogenous klotho upregulation, or recombinant Klotho supplementation promote neurogenesis. Increased klotho expression may preserve the NPCs’ niche, stimulate NPCs proliferation, increase the number of neuroblasts and immature neurons, and enhance dendritic arborization. The indiferentiated molecular markers are Nestin and glial fibrillary acidic protein (GFAP). The proliferation markers are SOX2 and KI67. Doublecortin (DCX) labels immature neurons, and NeuN labels mature neurons. Created in BioRender (https://BioRender.com/zn72n28).
Comments (0)