
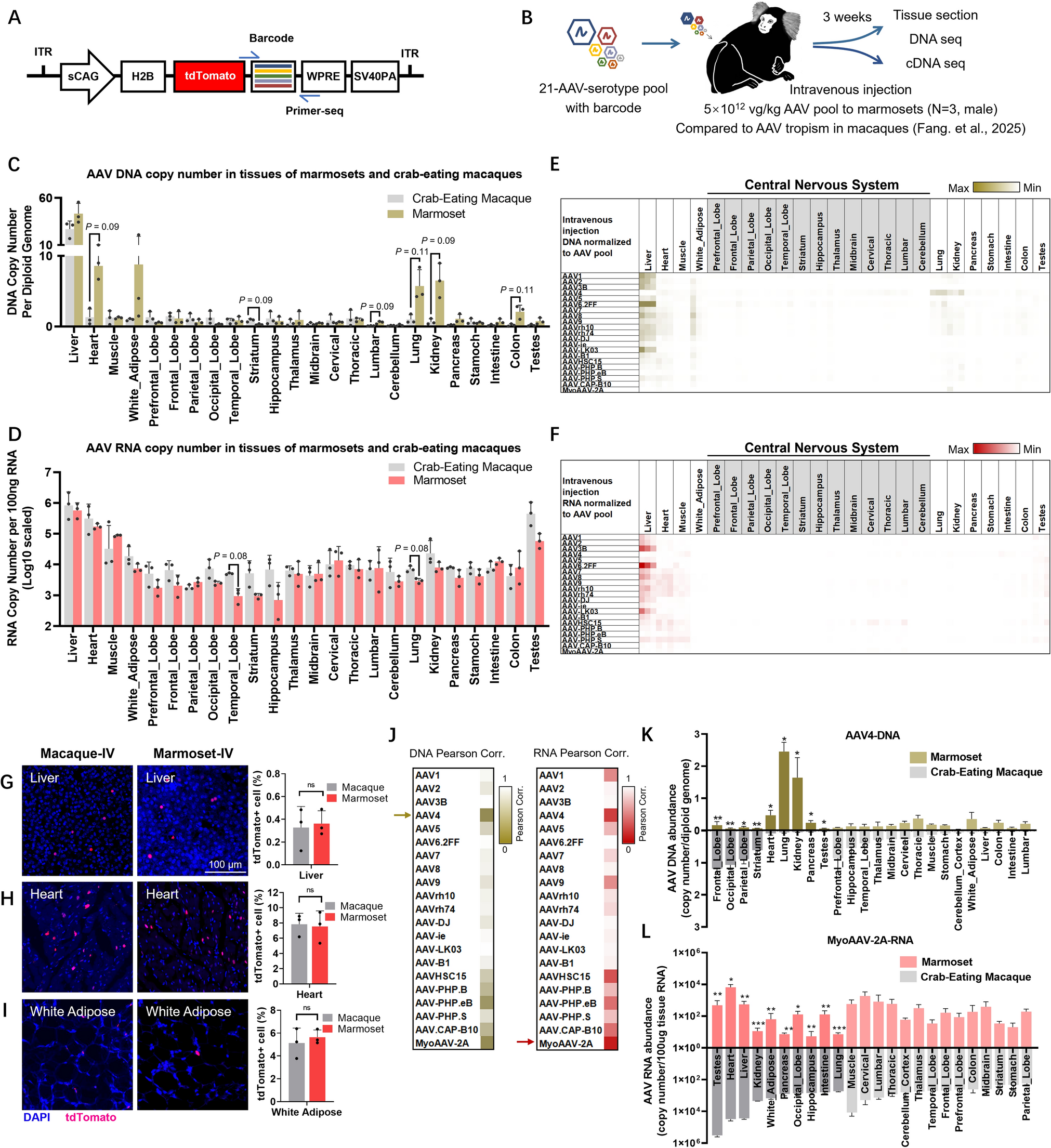
Remember me

To comprehensively investigate the cellular and molecular changes occurring during epileptogenesis, we first established a PILO-induced epilepsy model in rats. Following the intraperitoneal injection of PILO, the rat model exhibited the typical behavioral characteristics of the five seizure stages on the Racine scale [24]: Stage 1, mouth and facial movements; Stage 2, head nodding; Stage 3, forelimb clonus; Stage 4, rearing; and Stage 5, rearing and falling. We used video behavioral monitoring, electroencephalography (EEG), histological staining, and qRT-PCR to confirm the successful establishment of the epilepsy model.
Specifically, only seven rats were included in the chronic group after eight weeks of video behavioral monitoring (Table S1). The average latency period was 21.43 ± 7.39 d, with an average frequency of SRSs of 0.59 ± 0.93 times per day and an average SRS duration of 38.57 ± 10.42 s. None of the rats in the control group developed SE or SRSs. EEG recordings performed in the fourth week post-SE revealed interictal epileptic discharges and ictal spike rhythms during SRSs in epileptic rats, but not in control rats, using the same batch of animals employed for snRNA-seq (Fig. 1A). Histological analysis via Nissl staining demonstrated persistent neuronal death in the CA1 region of the hippocampus during epileptogenesis, with more pronounced neuronal loss in the acute phase. Additionally, a slight decrease in temporal cortex neurons and atrophy in the thalamus were observed, particularly during the chronic phase (Fig. 1B). These findings align with previous observations in TLE epilepsy models [15].
Fig. 1
Establishment of the pilocarpine-induced epilepsy rat model. A EEG recordings taken in the fourth week after pilocarpine injection show interictal epileptic discharges and spontaneous seizures. The blue arrows in the second episode highlight spike waves, in contrast to the control group (first episode). Epileptic rats exhibited spontaneous seizures characterized by rhythmic spike waves with a frequency >2 Hz and an amplitude three times the baseline (third episode). B Nissl staining was performed on the CA1 region of the hippocampus, the temporal cortex, and the thalamus in the control group and the three phases. Neuronal loss was persistent in the CA1 region throughout all phases and was particularly pronounced during the acute phase. The temporal cortex and thalamus also exhibited slight neuronal death, with notable atrophy observed in the thalamus, especially in the chronic phase. C The relative mRNA expression levels of Tnf-α (F value=4.221), Il-1β (F value=11.43), C3 (F value=7.043), C1qa (F value=11.36), vGlut1 (F value=12.61), and vGat (F value=9.853) in the hippocampus were compared across three phases with those in the control group (n=4–8 per group). The P-value was calculated using one-way ANOVA. * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001. N, control group; A, acute phase; L, latent phase; C, chronic phase. Astro-astrocytes, Micro-microglia, EX-excitatory neuron, IN-inhibitory neuron, OL-oligodendrocyte, OPC-oligodendrocyte precursor cell, Endo-endothelial cell.
At the molecular level, significant inflammatory activation was observed, with mRNA expression levels of inflammatory factors Tnf-α and Il-1β significantly increased in the acute phase (Fig. 1C). Complement activation was also noted, with mRNA levels of complement C3 and C1qa upregulated during epileptogenesis, particularly in the chronic phase (Fig. 1C). Furthermore, synaptic loss was observed, with decreased mRNA levels of the excitatory synapse marker vGlut1 and the inhibitory synapse marker vGat in the epilepsy group during epileptogenesis (Fig. 1C). These findings are consistent with previous studies [8, 41, 42]. Thus, we confirm that we have successfully established a rat TLE model via intraperitoneal PILO injection.
Single-Nucleus Transcriptomic Atlas of a Pilocarpine Rat Model of EpileptogenesisWith the established rat epilepsy model, we conducted a single-nucleus transcriptomic analysis. Single-nucleus sampling focused on four groups of rats: the control group (7 d after receiving an equal volume of saline as a substitute for PILO), the acute phase group (24 h post-SE), the latent phase group (7 d post-SE), and the chronic phase group (8 w post-SE) of epileptogenesis (Fig. 2A). Three individual rats were used as biological replicates for each group. For the twelve animals used for snRNA-Seq, the average dose of pilocarpine administered in the epilepsy group was 23.33±5.00 mg/kg, while the average saline dose in the control group was 23.33 ± 5.78 mg/kg. The total doses for the experimental and control rats averaged 34.33 ± 5.00 mg/kg and 34.33 ± 5.77 mg/kg, respectively. The latency to SE (time from initial injection to SE onset) averaged 31.78±4.13 min in the epilepsy group, consistent with previous studies [11] (Table S2).
Fig. 2
Single-nucleus transcriptomic atlas of the pilocarpine rat model during epileptogenesis. A The hippocampus, temporal cortex, and thalamus of rats from both the control group and the three phases were collected at defined time points. Three individual rats were used as biological replicates for each group, resulting in a total of 36 samples for snRNA-Seq analysis. Continuous 24-hour video recordings were conducted on all animals during the first seven days post-SE to ensure that none of the rats exhibited spontaneous seizures during the latent phase. Beginning on the 8th day after modeling, surviving animals were monitored via continuous video for 12 h daily over the next 7 weeks to detect SRSs. EEG recordings were conducted on a subset of animals from the same experimental batch used for snRNA-Seq in the fourth week and were excluded from subsequent experiments. B UMAP representation of 311,177 nuclei from 36 samples, showing nine clusters, each represented by a different color. C Dot plots displaying the marker genes for each cluster. D–F Changes in cell abundance in the temporal cortex during the acute (D) and chronic (E) phases, as well as in the thalamus during the latent phase (F), compared to the control group. Neighborhoods that overlap with the same cell population are grouped together and colored to indicate significant differential abundance (DA) (Spatial FDR < 0.1). Cell clusters with significantly decreased abundance are shown in red, while those with significantly increased abundance are shown in blue. N, control group; A, acute phase; L, latent phase; C, chronic phase. Astro-astrocytes, Micro-microglia, EX-excitatory neuron, IN-inhibitory neuron, OL-oligodendrocyte, OPC-oligodendrocyte precursor cell, Endo-endothelial cell.
Tissue samples were collected from three brain regions (the hippocampus, temporal cortex, and thalamus) of each rat, resulting in a total of 36 samples. SnRNA-seq was performed on these samples, and after quality control, we obtained a total of 311,177 single nuclei. These nuclei were annotated into nine cell types (Fig. 2B, C), including astrocytes, microglia, excitatory neurons (EXs), inhibitory neurons (INs), oligodendrocytes (OLs), ependymal cells, oligodendrocyte precursor cells (OPCs), fibroblasts, and endothelial cells. Detailed information on the rats used for snRNA-seq and cell types for each sample is provided in Table S2.
We further analyzed cell type abundance changes during epileptogenesis and observed notable alterations (Fig. 2D–F and Table S3). In the acute phase, the temporal cortex exhibited significant shifts in the abundance of various cell types, reflecting dynamic cellular activity (Fig. 2D). During the chronic phase, we identified a reduction in endothelial cells and astrocytes, accompanied by an increase in OLs and microglia in the temporal cortex (Fig. 2E). In the latent phase, the thalamus showed an increased abundance of microglia, a decreased abundance of OPCs and astrocytes, along with alterations in OLs (Fig. 2F). These findings underscore dynamic cellular responses across different brain regions during epileptogenesis, with the most prominent changes occurring in the temporal cortex during the acute phase.
Differentially Expressed Genes (DEGs) During EpileptogenesisBuilding on our investigation of dynamic cellular changes, we further explored the molecular and genetic alterations that occur during epileptogenesis. We compared the gene expression levels in each cell type of the epilepsy groups (across three brain regions and three phases) to those of the corresponding cell type in the control group, identifying a total of 2,449 DEGs in different brain regions during epileptogenesis (Fig. 3A–C, Table S4, and Fig. S1). The acute phase exhibited the highest number of DEGs across all brain regions (1,775), with the hippocampus showing the greatest number (1,007, compared with 947 in the temporal cortex and 627 in the thalamus) (Fig. 3A). This highlights that molecular changes in the hippocampus are most prominent during the onset of epileptogenesis. As epileptogenesis progressed to the latent phase, the thalamus exhibited the highest number of DEGs (789, compared with 40 in the hippocampus and 116 in the temporal cortex) (Fig. 3A, B). Notably, 707 of the 789 DEGs in the thalamus were upregulated. Finally, in the chronic phase, the number of DEGs in the thalamus decreased substantially compared to the latent phase (13 compared with 789), while the number of DEGs in the hippocampus increased compared to the latent phase, though still lower than in the acute phase (168 compared with 40 and 1,007, respectively) (Fig. 3A, B). These findings suggest that, similar to the acute phase, the hippocampus may also serve as the primary site of molecular changes during the chronic phase of epileptogenesis.
Fig. 3
Differentially expressed genes (DEGs) during epileptogenesis. A Bar plot showing the number of upregulated, downregulated, and total DEGs in the hippocampus, temporal cortex, and thalamus across the three phases. Upregulated genes were defined as those with adjusted P values < 0.05 and log2fold changes > 0.25, while downregulated genes were defined as those with adjusted P values < 0.05 and log2fold changes < −0.25. B Venn diagrams (http://www.ehbio.com/test/venn) illustrating the similarities and differences among DEGs identified across the three phases and brain regions. A total of 46 genes were differentially expressed in all three phases, while 249 genes were differentially expressed across all three regions. C Region-specific genes and latent phase-specific (547) genes were further analyzed. The thalamus exhibited the greatest number of common genes (495) with latent phase-specific genes, compared to the hippocampus (12) and temporal cortex (17). D Functional enrichment analysis of the 495 common genes shared between the thalamus and latent phase-specific genes. E Functional enrichment analysis of the 249 genes that were differentially expressed across all three brain regions as well as the 46 genes that were differentially expressed in all three phases.
Furthermore, we compared the similarities and differences between DEGs identified across the three phases and brain regions (Fig. 3B). The acute phase had the greatest number of unique DEGs (1,411), whereas 46 genes were differentially expressed in all three phases. These genes could serve as molecular markers for identifying epileptogenesis. Genes with phase-specific differential expression could also act as molecular signatures distinguishing different disease stages. Additionally, we identified 249 genes that were differentially expressed across all three brain regions, with 435, 314, and 871 genes exclusively expressed in the hippocampus, temporal cortex, and thalamus, respectively (Fig. 3B, C). Region-specific analysis revealed that among 547 genes specific to the latent phase, the thalamus had the largest subset (495), compared with the hippocampus (12) and temporal cortex (17) (Fig. 3C), further implicating the thalamus as a major site of molecular changes during the latent phase. Functional enrichment analysis of these 495 genes (Fig. 3D) identified enriched pathways, such as regulation of membrane potential, trans-synaptic signaling, and synapse organization. Taken together, these findings suggest that the thalamus may play a significant role in synaptic remodeling during the latent phase.
In addition, to elucidate the roles of the 46 genes differentially expressed across all phases and the 249 genes expressed across all regions, we performed functional enrichment analysis on these genes (Fig. 3E). The 249 genes expressed across all brain regions were enriched in pathways related to glial differentiation, neuronal remodeling, and synaptic remodeling. Several of these genes, including Gfap [43], Stat3, Atp1b2, Eef2k [44], S100b [45], Spp1, Agt, Cd14, and Igfbp [46], have been implicated in human epilepsy or animal models. Meanwhile, the 46 genes altered across all three phases were associated with metabolism, biosynthesis, and cell-cell adhesion. Some of these genes, such as Cnn3 [47], Cst3 [48], Gjb6 [49], Hspb1 [50], and Kcnip2 [51], have been previously reported in epilepsy studies.
In summary, these findings suggest that glial differentiation, neuronal remodeling, and synaptic remodeling processes occur in the hippocampus, temporal cortex, and thalamus during epileptogenesis, while metabolic and biosynthetic processes play roles at different stages of the disease.
Critical Genes Involved in EpileptogenesisTo reveal comprehensive and specific molecular changes in the development of epileptogenesis, we further identified critical genes that may be involved in this process. By comparing the three phases with the control, we calculated the number of cell types in which the identified DEGs showed differential expression and listed genes that exhibited differential expression across most cell types. In the acute phase, genes including Spp1, Hspa1a, Timp1, Hspb1, Socs3, Stat3, Htra1, Sbno2, Gpnmb, and Vim were differentially expressed across most cell types (Fig. 4A), with specific cell types detailed in Fig. 4D. Among these key genes, those related to inflammation, such as Spp1, Socs3, Stat3, Hsps, and Sbno2, may play roles in sensing stress and transmitting stress signals to downstream genes, while Htra1, Gpnmb, Timp1, and Vim reflect dramatic structural changes in various cell types, embodying the two key features of molecular alterations in the acute phase. Compared with the acute phase, fewer genes were differentially expressed in multiple cell types during the latent and chronic phases. In the latent phase, Nrg3, Kcnip4, Nrxn3, Rims1, Ptprd, Xkr4, Syt1, Nav3, Ralyl, and Il1rapl1 were the most frequently identified DEGs across cell types (Fig. 4B, E), primarily expressed in the thalamus. These genes are involved in electrical signal generation and conduction (Kcnip4, Syt1, and Nav3), as well as cell migration and proliferation (Nrg3, Ptprd, Nrxn3, Ralyl, and Rims1), which may relate to synaptic remodeling. Finally, in the chronic phase, the key genes included Ctss, Gfap, Cd74, Trh, Gpd1, Npy, Ptgs2, Slc6a1, Tac3, and Cnp (Fig. 4C, F), mainly expressed in the hippocampus. These genes are associated with immune regulation (Ctss, Cd74, Gfap, and Ptgs2), neural regulation (Npy, Slc6a1, and Cnp), and metabolism regulation (Gpd1, Trh, and Tac3). In conclusion, during the acute and chronic phases, gene expression differences were most evident across cell types in the hippocampus, whereas differences in some genes were less pronounced in cells of the temporal cortex and thalamus. However, during the latent phase, expression differences were primarily observed in thalamic cell types, suggesting significant genetic alterations in the thalamus during this period.
Fig. 4
Key differentially expressed genes (DEGs) and four major gene clusters involved in epileptogenesis. The top 10 DEGs in nine cell types across the acute (A), latent (L), and chronic phases (C) compared with those in the control group (N) are presented in bar plots A, B, and C. The number of cell types is labeled on the bar plots, with detailed cell types illustrated in D, E, and F, respectively. G The number of common genes identified in each cell type across the three brain regions and three phases. H Time-trend clustering of DEGs. Four cluster categories were identified based on their gene expression patterns among the control, acute, latent, and chronic phases: Cluster 1, genes significantly upregulated in the chronic phase. Cluster 2: Genes significantly upregulated in the acute phase. Cluster 3: Genes significantly downregulated in the acute phase. Cluster 4: Genes significantly upregulated in the latent phase. I Gene functional enrichment analysis of the four clusters highlights their respective biological roles during different phases of epileptogenesis.
We further analyzed changes in DEGs across various cell types. Specifically, for each cell type (OLs and OPCs grouped together) in different brain regions, we compared them to the control group across the three phases of epileptogenesis to identify DEGs. Our focus was on genes that consistently showed differential expression across all three phases (Fig. S2 and Table S5) within each cell type, as well as those shared across different brain regions (Fig. S3 and Table S5), as these genes are more likely to be strongly related to epileptogenesis. The total number of such common genes is summarized in Fig. 4G, with specific genes detailed in Table S5. Gene function enrichment analysis was performed on the common genes across the three brain regions (Fig. S4A) and the three phases (Fig. S4B). Genes commonly altered across brain regions were primarily associated with tissue remodeling, cellular signaling, gliogenesis, immune responses, metabolism, and homeostasis. In contrast, genes commonly altered across phases were associated with cell migration and development, glial cell differentiation, and metabolic responses. Specifically, we identified three genes—Gjb6, which was downregulated across the three brain regions and phases of epileptogenesis in astrocytes; Ankrd55, which was downregulated; and Wapl, which was upregulated across the three brain regions and phases of epileptogenesis in microglia (Fig. 4G, Fig. S5, and Table S5). Gjb6 encodes Connexin 30 (Cx30), a protein in the connexin family that forms gap junctions between cells. Previous research showed that Cx30 was overexpressed in the hippocampus at both the transcriptional and translational/posttranslational levels at the beginning of kindling epileptogenesis. This overexpression remained at the mRNA level (but not at the protein level) after focal seizures were acquired, but was significantly downregulated in epileptic animals [49]. This suggests that Cx30 plays a crucial role in epileptogenesis. Ankrd55 is associated with some autoimmune disorders like multiple sclerosis and rheumatoid arthritis [52], while Wapl regulates cohesin's release from chromatin by opening a distinct DNA exit gate [53]. Although the roles of Ankrd55 and Wapl in microglia during epileptogenesis are not well understood, our results suggest their potential involvement in this process.
Additionally, we observed that EXs and INs shared few common genes across regions and phases, indicating significant gene expression variability in neurons across different phases and regions during epileptogenesis. In contrast, glial cells, including astrocytes, microglia, and OLs/OPCs, exhibited many shared genes across all three regions, suggesting glial cells may drive similar processes across brain regions. Further analysis of five common genes expressed in glial cells—Hspa1a, Hspb1, Spp1, Stat3, and Vim (Table S5)—revealed their involvement in inflammation and cytokine signaling pathways, indicating that glial cells may mediate inflammatory responses across all brain regions during epileptogenesis.
Different Gene Clusters Revealed Functional Changes During EpileptogenesisTo better understand the functions of the 2,449 DEGs identified during epileptogenesis, we classified all DEGs into four gene clusters (Fig. 4H and Table S6) based on their gene expression changes across the control group, acute phase, latent phase, and chronic phase. These clusters included: (1) genes significantly upregulated in the chronic phase (Cluster 1); (2) genes significantly upregulated in the acute phase (Cluster 2); (3) genes significantly downregulated in the acute phase and subsequent phases (Cluster 3); and (4) genes significantly upregulated in the latent phase (Cluster 4). To elucidate the functions of specific genes within each cluster, functional enrichment analysis was conducted (Fig. 4I). Genes significantly downregulated in the acute phase (Cluster 3) were primarily associated with various cellular metabolic processes, as well as cell growth and development. In contrast, genes significantly upregulated in the acute phase (Cluster 2) were related to cell regulation and adhesion, and the apoptotic signaling pathway. These findings indicate that during the acute phase, metabolic synthesis processes, neuronal development, and synaptic maturation are impaired, accompanied by neuronal apoptosis and neuroinflammation. Genes upregulated in the latent phase (Cluster 4) were enriched in pathways associated with synapses and membrane potential, highlighting the latent phase as a critical period for synaptic and neural network remodeling during epileptogenesis. Finally, Cluster 1, which included genes upregulated in the chronic phase, showed enrichment in pathways related to immune processes.
Together, these four distinct clusters of DEGs reveal the major molecular changes occurring across the different phases of epileptogenesis, offering insight into the phase-specific mechanisms driving the disease.
Cell-Cell Interaction Dynamics Indicate Strong Intercellular Communication During the Latent PhaseCell-cell interactions play a crucial role in regulating various biological processes [54]. Accordingly, we analyzed the interactions among different cell types during epileptogenesis (Fig. 5 and Table S7). Regardless of the quantity (Fig. 5A) or intensity (Fig. 5B), cell interactions were found to be strongest during the latent phase—a trend that remained consistent across all three brain regions. In contrast, the most pronounced changes in gene expression occurred during the acute phase, particularly in the hippocampus, where cell interactions were at their lowest compared to the control group. However, interactions in the temporal cortex and thalamus showed relative increases compared to the control group during the acute phase.
Fig. 5
Cell-cell interaction dynamics during epileptogenesis. Number (A) and intensity (B) of cell interactions across the three brain regions during epileptogenesis. C, D, and E show the number of common and specific cell interaction pathways in the hippocampus (C), temporal cortex (D), and thalamus (E) during different phases. The cell interaction pathways used in the hippocampus, temporal cortex, and thalamus across different phases are detailed in F, G, and H, respectively, with the specific pathways labeled in blue.
We then examined the number of cell interaction pathways and their usage across different phases and brain regions (Fig. 5C–H). Specifically, we focused on the unique signaling pathways in the three brain regions at different phases, as highlighted in Fig. 5F–H. To further clarify the specific cell types involved in these unique signaling pathways, we used heatmaps to visualize cell communications among different cell types across the three brain regions during epileptogenesis (Figs. S6–S8). During the acute phase, four exclusive signaling pathways were utilized in the hippocampus: TWEAK, BRADYKININ, SELE, and VISFATIN (Fig. 5C, F). Specifically, the TWEAK and VISFATIN signaling originated from endothelial cells and targeted astrocytes. SELE signaling was directed from endothelial cells to EXs, while BRADYKININ signaling was transmitted from fibroblasts to endothelial cells (Fig. S6). In the temporal cortex, the exclusive pathways included SPP1, SELE, IGFBP, VISFATIN, TULP, TWEAK, and EGF (Fig. 5D, G). Most of these signaling pathways were transmitted from endothelial cells and astrocytes to EXs and astrocytes (Fig. S7). In the thalamus, SPP1, TULP, and VISFATIN were identified as exclusive pathways (Fig. 5E, H). SPP1 originated from EXs and targeted astrocytes and endothelial cells, while TULP and VISFATIN were directed from endothelial cells to OLs and other endothelial cells (Fig. S8). During the latent phase, the hippocampus and thalamus each exhibited three distinct cell interaction signaling pathways. In the hippocampus, these included CD45, OPIOID, and CSPG4 (Fig. 5C, F), with CSPG4 and OPIOID originating from endothelial cells to INs, and CD45 from microglia to fibroblasts (Fig. S6). In the thalamus, the exclusive pathways were EPHB, NETRIN, and EPHA (Fig. 5E, H), all originating from endothelial cells and EXs, and targeting EXs (Fig. S8). In the chronic phase, only the thalamus exhibited two specific pathways: RBP4 and HISTAMINE (Fig. 5E, H). RBP4 originated from fibroblasts and targeted endothelial cells, while HISTAMINE was directed from ependymal cells to EXs (Fig. S8).
Furthermore, we summarized the number of cell communications as well as the dominant senders (sources) and receivers (targets) among different cell types in the three brain regions during epileptogenesis (Fig. S9, 10). Notably, the strongest communications were observed between EXs and INs, with EXs consistently serving as the primary input-output hub for interactive signals. Overall, cell interactions among most cell types were enhanced during the latent phase across the three brain regions. The enhancement of intercellular communication is a key feature of the latent phase and could serve as a potential target for future intervention strategies.
Two Novel Astrocyte Clusters and the Specific EX-Astro C3-IN Pathway Found in the Acute PhaseAccumulating evidence indicates that astrocytes contribute to the development and progression of hyperexcitability in epilepsy due to dysfunctions in gliotransmission, cell metabolism, and immunity [55]. In our single-nucleus analysis of the epileptic brain, we observed notable changes in the number of DEGs in astrocytes during epileptogenesis. Therefore, we further performed subcluster analysis of astrocytes and identified a total of 13 subpopulations (Fig. 6A). Further investigation into astrocytes revealed two distinct clusters (Cluster 3 and Cluster 9) that were significantly increased in the acute phase compared to those in the control group (Fig. 6B and Fig. S11). These clusters remained less abundant in the latent and chronic phases. Notably, they were present in all three brain regions, with the highest abundance in the hippocampus (Fig. S12).
Fig. 6
Two novel astrocyte clusters found in the acute phase of epileptogenesis. A Subcluster analysis of astrocytes identified a total of 13 subpopulations. B Two specific subpopulations, Cluster 3 and Cluster 9, were significantly increased during the acute phase compared to the control group. C CytoTRACE was used to predict transformation potential, with red indicating the starting point of transformation and blue indicating the endpoint. Both Clusters 3 and 9 were positioned at the end of the astrocyte trajectory, especially Cluster 9. D Trajectories of astrocytes in the acute phase were modeled using Monocle 3, with red lines emphasizing the transformation trajectories toward Cluster 3 and Cluster 9. E Cluster analysis of driver genes along the transformation paths to Cluster 3 and Cluster 9 was conducted, and the elbow method determined two optimal subclusters. F Heatmaps illustrated dynamic changes in driver genes along the pseudo-time trajectory toward Cluster 3 and Cluster 9. Additionally, functional enrichment analysis results for each clustering group were included. G, H Gene functional enrichment analysis of genes in Cluster 3 (G) and Cluster 9 (H).
Analysis of their formation process (Fig. 6C) revealed that these clusters are well-developed derivatives of astrocytes, as supported by pseudo-temporal cell trajectory inference (Fig. 6D), which positioned them at the terminal end of the astrocyte trajectory. Potential driver genes associated with their formation were identified through trajectory analysis (Fig. 6E and Table S8). Notable differences in driver genes for the two clusters suggest distinct formation processes (Fig. 6E, F). Cluster 3 showed enrichment for genes related to oxidative phosphorylation, oxidative stress response, cell migration, and synapse organization (Fig. 6F), while Cluster 9 was enriched with genes involved in the heat stress response, calcium ion response, and positive regulation of apoptosis (Fig. 6F). Functional enrichment analysis of genes uniquely expressed in these clusters (Fig. 6G, H) revealed pathways related to glial cell proliferation, indicative of an active proliferative state. Additionally, pathways responding to environmental factors such as external stimuli, hypoxia, polypeptides, and metal ions were enriched, suggesting a role in intercellular signaling and communication. Enrichment in synaptic transmission pathways further pointed to potential roles in regulating neuronal communication.
We next examined the cell-cell interactions involving these newly identified clusters to elucidate their functional mechanisms. Focusing on the acute-phase hippocampus, where these clusters were most prominent (Fig. S12), we compared cell-cell interactions between the acute-phase hippocampus group and the control group (Fig. 7A, B, and Table S9). While overall cell interactions decreased in the acute phase, the strength of interactions between astrocytes and several other cell types increased. Notably, several pathways absent in the control group, such as SPP1, COLLAGEN, and NT, emerged in the acute phase. Among these, SPP1-mediated interactions displayed the highest input and output intensities for astrocytes (Fig. 7C). Investigation of the SPP1 pathway across cell types revealed that EXs were the primary senders of SPP1 signals, while astrocyte C3 was the main receiver (Fig. 7D). Interestingly, astrocyte C3 also emerged as the most influenced cells, highlighting a specific influence of EXs on astrocyte C3 via the SPP1 pathway during the acute phase. To explore whether astrocyte C3 influenced other cell types, we analyzed additional acute-phase-specific pathways. The EGF pathway was found to be specifically active, with astrocytes—particularly Cluster 3—serving as the main source of EGF signals, and INs being the primary receivers (Fig. 7E). Based on these findings, we propose that during the acute phase, EXs influence astrocyte C3 through the SPP1 pathway, which subsequently affects INs via the EGF pathway (Fig. 7F). We used NicheNet to infer ligand-to-target signaling pathways based on a mouse dataset. Key genes in the SPP1 pathway included Spp1, Cd44, Itgav, Itgb1, Cd40, Ctnnb1, Egr1, Jun, Jund, Myc, Nfkb1, Smad3, Stat3, Chuk, Map3k1, Map3k14, Mapk1, Mapk8, and Traf3. Enrichment analysis revealed functions such as the MAPK cascade, positive regulation of cell migration, and focal adhesion (Fig. S13A). Similarly, key genes in the EGF pathway included Hbegf, Erbb4, Egfr, Stat1, Stat5a, Trp53, and Ptk2, with enrichment functions such as the ErbB signaling pathway and receptor signaling via JAK-STAT (Fig. S13B).
Fig. 7
Cell-cell interactions in the hippocampus during the acute phase of epileptogenesis. A Differences in the number of cell-cell interactions in hippocampal cells between the acute phase and the control group are shown, with increases highlighted in red and decreases in blue. B Differences in the intensity of cell-cell interactions between the acute phase and the control group are visualized, with increases in red and decreases in blue. C Differential interaction pathways of astrocytes in the hippocampus between the acute phase and the control group. The SPP1 pathway is identified as a specific input-output pathway. D SPP1 pathway network across different cell types. EXs were the primary senders of SPP1 signals, while astrocyte Cluster 3 served as the main receiver. E EGF pathway network across different cell types. Astrocytes, particularly Cluster 3, were the primary source of EGF signals, with INs being the main receivers. F Schematic diagram of the EX-Astro C3-IN pathway mediated by the SPP1 and EGF pathways in the hippocampus during the acute phase. EXs influence astrocyte C3 via the SPP1 pathway, while C3 further influences INs through the EGF pathway. Critical genes and enrichment functions are listed. The schematic diagram was created using BioRender.com. Astro-astrocytes, Micro-microglia, EX-excitatory n
Comments (0)