
Remember me



Previously, three regions of potential sequence homology between Corolla and SYP-4 were reported (Collins et al. 2014). Upon further investigation of the protein and its sequence, we found additional similarities to SYP-4 in C. elegans and SIX6OS1, which were recently suggested to be orthologs (Köhler et al. 2025; Neves et al. 2025). The C-terminus of both proteins is rich in phenylalanine compared to the entire protein sequence, and both proteins feature LC3-interacting region (LIR) motifs. Neves et al. (Neves et al. 2025) and (Köhler et al. 2025) report that specific perturbations within the SYP-4 C-terminus significantly disrupt the regulation of crossover formation. We therefore investigated whether Drosophila Corolla is a SIX6OS1 ortholog and found that the C-terminus was indeed enriched with phenylalanine. In D. melanogaster, 10% of the last 89 amino acids of Corolla were phenylalanine, compared to 4% in the entire protein sequence. Similarly, in D. mauritiana Corolla 9% of the last 89 amino acids were phenylalanine compared to 4% in the entire amino acid sequence. Additionally, we found two LIR motifs in Corolla protein sequences from D. melanogaster and its D. mauritiana ortholog using the Eukaryotic Linear Motif (ELM) resource for functional sites in proteins (Supplementary Fig. 1) (Kumar et al. 2024). One of these motifs is shorter by two amino acids in D. melanogaster, which means that through the gene replacement, this motif was extended, and it additionally contains 3 amino acid substitutions. One of these is an amino acid substitution of a phenylalanine with an aspartic acid at the aligned residue 351 (residue 349 in the D. melanogaster protein sequence). Phenylalanines in LIR motifs were shown to have a special functional importance in SYP-4 (Neves et al. 2025) but the importance of this one single phenylalanine is unknown. In combination with the previously highlighted homology regions between SYP-4 and Corolla (Collins et al. 2014) it is likely that Corolla is the Drosophila ortholog of SIX6OS1.
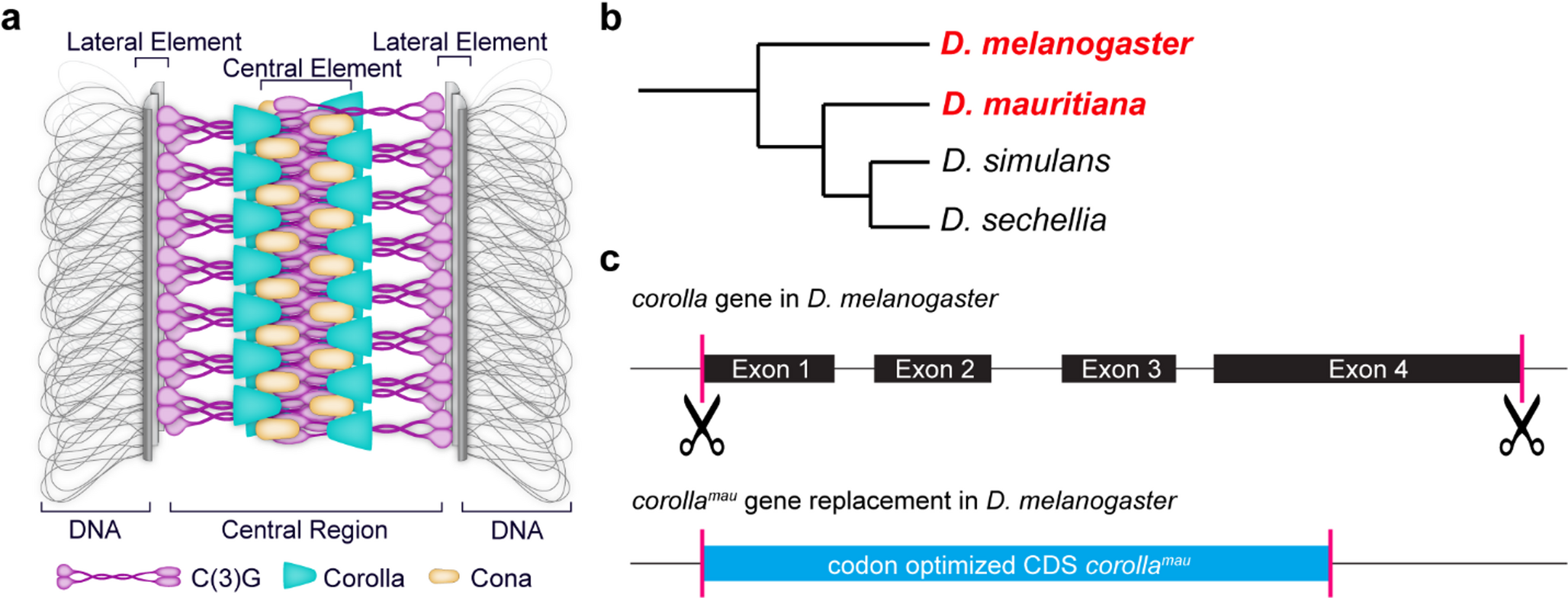
Replacing the corolla gene in D. melanogaster with its ortholog from D. mauritiana to create a hypomorphic alleleTo circumvent the problem of identifying conserved amino acid motifs that we could target for the creation of a hypomorphic allele, we chose to replace the D. melanogaster gene encoding the central region protein, Corolla, with that of a close relative. The closest relatives of D. melanogaster are D. mauritiana, D. simulans, and D. sechellia (Fig. 1b). Previous studies of recombination in D. mauritiana and D. simulans revealed that the total map length of D. mauritiana was 1.36 (Hawley et al. 2025) to 1.8 (True et al. 1996) times more compared to D. melanogaster, and the total map length of D. simulans was 1.3 times more compared to D. melanogaster (True et al. 1996). Both species exhibit more recombination in the regions proximal to the centromere than D. melanogaster (True et al. 1996; Hawley et al. 2025), but the strongest effect was seen in D. mauritiana. Therefore, we chose D. mauritiana as the donor for the replacement of the endogenous corolla gene in D. melanogaster, as it would allow us more easily to understand whether Corolla is directly involved in these differences of crossover patterning.
The amino acid sequence of Corolla from D. mauritiana shares 86.7% sequence identity with D. melanogaster Corolla (Supplementary Fig. 2 and Supplementary file 1). In total, there are 74 amino acid differences including a two amino acid gap (Supplementary Fig. 2 and Supplementary file 1). These amino acid differences have a similarity of 4.7%, meaning that a substituted amino acid is of similar chemical type. The DeepCoil2 program (Zimmermann et al. 2018; Ludwiczak et al. 2019; Gabler et al. 2020) indicates that Corolla from D. mauritiana has an overall higher probability for coiled-coil formation within the first 200 amino acids of the protein than D. melanogaster (Supplementary Figs. 3a-b). To form coiled-coils, a heptad repeat of seven amino acids must be present, usually denoted as a-b-c-d-e-f-g (McLachlan and Stewart 1975). The positions a and d within that heptad repeat are typically hydrophobic amino acids while positions e and g are typically charged or polar amino acids (McLachlan and Stewart 1975; Parry 1982; Lupas et al. 1991). The higher probability of coiled-coil formation in Corolla from D. mauritiana is likely because DeepCoil2 finds more residues fitting the above-described a and d positions of heptad repeats within the first 200 amino acids (Supplementary Figs. 3a-b). Importantly, the length of the region with coiled-coil probability is predicted to be of the same length within the first 200 amino acids of the proteins, and the arrangement of individually predicted coiled-coil domains is largely similar in spacing (Supplementary Figs. 3a-b). This is reminiscent of the findings by Kursel et al. which showed that within nematodes the length and location of coiled-coil domains is highly conserved as well as the overall protein lengths of orthologous SC proteins (Kursel et al. 2021). Indeed, the overall length of the Corolla protein from both Drosophila species is strikingly similar with 554 amino acids in D. melanogaster and 556 amino acids in D. mauritiana (Supplementary Figs. 2–3 and supplementary file 1). In contrast, AlphaFold3 predictions of a Corolla homodimer from D. melanogaster shows a distinctly longer coiled coil (approx. 200 A) than from D. mauritiana (approx. 300 A) (Supplementary Fig. 4). It is unclear how exactly this may affect Corolla and its interaction partners.
In addition to a higher probability of coiled-coil formation, we also find that Corolla from D. mauritiana has a slightly larger intrinsically disordered region (IDR), as well as a small additional IDR within the C-terminus of the protein that is not found in Corolla from D. melanogaster according to an InterProScan (Supplementary Figs. 3C-D) (Blum et al. 2025). Specifically, the IDR in Corolla from D. melanogaster spans amino acids 308–432 while the first IDR in Corolla from D. mauritiana spans amino acids 305–462, and the second small IDR spans amino acids 476–495 (Supplementary Fig. 3c-d). IDRs have been established as important factors in the function of SC proteins. Especially in SYP-4 of C. elegans, it is the IDR that is important for the regulation of crossovers and maintenance of biophysical characteristics of the SC (Köhler et al. 2025; Neves et al. 2025). Importantly, IDRs and coiled-coil domains are implicated in promoting phase-phase separation through multivalent interactions (Zhang et al. 2020; Kursel et al. 2021).
In light of the differences in protein architecture described above and the high possibility of Corolla being a SIX6OS1 ortholog, we hypothesized that replacing D. melanogaster Corolla with its ortholog from D. mauritiana would allow for synapsis but could impact recombination. To generate D. melanogaster flies expressing the D. mauritiana Corolla protein, we used CRISPR/Cas9 to excise corolla exons 1–4 and inserted the codon-optimized coding sequence (CDS) of D. mauritiana corolla (Fig. 1c). We wanted to insert the CDS in the endogenous corolla locus so that regulatory elements of corolla gene expression could act on the gene expression of the replacement. We also chose not to insert the introns from D. mauritiana to avoid problems caused by foreign intron sequences. The resulting allele, corollamau, was then subjected to detailed phenotypic analysis.
corolla mau females exhibit early SC disassemblyUpon completion of the gene replacement, we first asked how this affected SC assembly, maintenance and disassembly. Within the germarium of D. melanogaster, meiotic progression can be followed spatially (Fig. 2a). Using structured illumination microscopy (SIM) on corollamau mutant germaria maintained at 25 °C, we observed that the SC assembled but was not maintained beyond early pachytene (Fig. 2b). Specifically, compared with wild-type controls (= corolla+) we observed early SC disassembly in 4/37 nuclei in early pachytene, 14/20 nuclei in early-mid pachytene, and 3/3 of nuclei in mid pachytene (Supplementary Fig. 5). To test whether the SC width was affected in corollamau mutants we turned to stimulated emission depletion (STED) microscopy, which offers higher resolution than SIM. We used a C(3)G antibody that recognizes a region in the C-terminal globular domain of C(3)G (Anderson et al. 2005). As the C-terminal region of C(3)G is localized in proximity to the lateral elements (Anderson et al. 2005; Cahoon et al. 2017), the fluorescence signal should clearly resolve into two tracks. The two tracks of C(3)G in both corolla+ and corollamau mutants were clearly visible in the STED images (Fig. 2d). We found that the SC width in corollamau germaria was not significantly different (124 ± 16 nm) compared to the corolla+ SC (124 ± 9 nm) as determined with a t test (p-value = 0.95) (Fig. 2c). STED microscopy did reveal that only 14% of SCs in early pachytene were full-length while 64% of SCs already showed mild fragmentation, and 21% of SCs show high levels of fragmentation (Supplementary Fig. 6). By early-mid pachytene, 70% of SCs were mildly fragmented while the remaining 30% were highly fragmented (Supplementary Fig. 6). The fact that fragmentation was apparent with STED but not SIM is likely due to the above-mentioned differences in the resolution of these microscopy techniques. Combined, these results show that that the replacement of Corolla from D. melanogaster with its ortholog from D. mauritiana allowed for the assembly of the SC, but that it could not be maintained beyond early pachytene, when SC fragmentation was already apparent.
Fig. 2
The corollamau gene replacement causes SC maintenance defects. (a) Schematic of the D. melanogaster germarium. Germline stem cells are found in the premeiotic region 1. These proliferate to a 16-cell cyst at region 2 A where two nuclei enter meiosis and assemble the SC in early pachytene and the remaining nuclei become nurse cells. As these cysts migrate along the germarium, they enter early-mid pachytene in region 2B where we still see two meiotic pro-oocyte nuclei. At mid pachytene, region 3, only one meiotic cell remains, the oocyte. The full-length SC is clearly visible from early pachytene to mid pachytene. (b) Immunofluorescence staining of the corolla+ and corollamau SC. In early pachytene, SC threads of both C(3)G (magenta) and Corolla (cyan) are present in corolla+ and corollamau. In early-mid pachytene, SC threads of C(3)G and Corolla are visible for corolla+ and corollamau reared at 18 °C. At the same stage, in corollamau reared at 25 °C the SC is visibly disassembling (a) or has already disassembled (b). In mid pachytene, SC threads are visible in corolla+ and corollamau reared at 18 °C, but the SC in corollamau flies reared at 25 °C is in the process of disassembling. Polycomplexes are visible in corollamau nuclei regardless of rearing temperature (marked by arrowheads). Scale bars represent 2 μm length. (c) Relative fluorescence intensity of the two C-terminal C(3)G tracks over distance in corolla+ and corollamau reared at 25 °C. The SC widths between corolla+ and corollamau are not statistically significantly different. (d) Deconvoluted maximum projection of the SC in corolla+ and corollamau imaged with STED microscopy. The two tracks of the C(3)G C-terminal antibody staining are visible. In corollamau, SC mild fragmentation is visible. Scale bars represent 2 μm length
SC-dependent centromere clustering in corolla mau mutantsIn D. melanogaster, the SC at the centromeres is different from the chromosome arms. Electron microscopy studies revealed that the central element has a less definite morphology and the lateral elements are surrounded by highly condensed chromatin making it more difficult to distinguish them from each other (Carpenter 1975). In addition, at the beginning of prophase I the centromeres cluster into one or two masses (Takeo et al. 2011). This process is called centromere clustering and requires the presence of C(3)G, Corolla, and Cona (Takeo et al. 2011; Collins et al. 2014). Due to the observed early disassembly and SC fragmentation, we questioned whether centromere clustering would be affected in corollamau flies. To analyze this, we stained germaria with an antibody against CID (Centromere Identifier, the Drosophila equivalent of CENP-A) and scored the number of clusters per nucleus (Supplementary Fig. 7). In early pachytene, the average number of CID clusters was 1.9 ± 0.8 for corolla+ and 1.8 ± 0.7 for corollamau (Supplementary Fig. 7). In early-mid pachytene, the average number of CID clusters was 1.9 ± 0.6 for corolla+ and 2.1 ± 0.8 for corollamau flies (Supplementary Fig. 7). Lastly, in mid pachytene, the average number of CID clusters was 1.8 ± 0.5 for corolla+ and 2.1 ± 0.7 for corollamau females (Supplementary Fig. 7). P values determined with an unpaired t-test indicated no statistical significance in any of these cases (Supplementary Fig. 7). Therefore, we can conclude that SC-dependent centromere clustering is not affected in corollamau mutants (Supplementary Fig. 7). This shows that while Corolla from D. mauritiana causes defects with SC maintenance, other functions of Corolla like centromere clustering can be successfully completed.
Polycomplexes in corolla mau flies do not consist of all SC protein componentsIn addition to the assembly of the tripartite SC that is not properly maintained, we noted the occurrence of polycomplexes (PC) in corollamau flies (Fig. 2b). PCs are structures consisting of SC proteins that assemble into repetitive structures that resemble the fine ultrastructure of the canonical SC (reviewed in (Hughes and Hawley 2020). While these are most often seen in mutant backgrounds that perturb the SC, well-studied examples of PCs include those induced by increased temperature in wild-type organisms (Bilgir et al. 2013; Rog et al. 2017). The spontaneous occurrence of PCs under standard conditions has also been reported, but it is not a common phenomenon (Anderson et al. 2005; Hughes and Hawley 2020). In corollamau mutant germaria, 1/37 pro-oocytes at early pachytene, 15/18 pro-oocytes at early-mid pachytene, and 3/3 oocytes at mid pachytene contained PCs, which were not observed in corolla+ (Fig. 2b and Supplementary Fig. 8a). Importantly, we found that 61% of PCs in corollamau flies lacked the SC component C(3)G (Fig. 2b and Supplementary Figs. 8a-b). To our knowledge, PCs in flies usually consist of all SC central region proteins (Hughes and Hawley 2020), which would make this the first observation of PCs lacking a central region component. The PCs are torroidal and appear hollow (Fig. 2b and Supplementary Figs. 8b-c). With this shape, the PCs are reminiscent of those seen in the c(3)gCdel mutants reported by Jeffress et al. (Jeffress et al. 2007), as well as sinaA4/sinaDf mutants reported by Hughes et al. (Hughes et al. 2019). However, all previously reported PCs contain C(3)G. To verify that the PCs were localized within the nucleus, we co-stained for the nuclear envelope protein Lamin and found that they were indeed nuclear (Supplementary Fig. 8c).
To summarize, we found that the replacement of D. melanogaster Corolla with its ortholog from D. mauritiana results in abnormal PC formation. Some of these PCs do not contain the transverse filament protein C(3)G, something that has never been observed before in flies. PCs may form due to the early disassembly of the SC, whereas the PCs lacking C(3)G may indicate a faulty interaction between C(3)G from D. melanogaster and Corolla from D. mauritiana.
The corolla mau SC maintenance defect is temperature-sensitiveDue to the occurrence of PCs, and especially those without the transverse filament protein C(3)G, we wondered whether the observed SC disassembly was caused by an unstable interaction between D. mauritiana Corolla and the native D. melanogaster proteins Cona and C(3)G. Corolla and Cona have been previously shown to interact with each other in a yeast two-hybrid assay (Collins et al. 2014). Importantly, it is still unclear how these two proteins interact with C(3)G.
Previous studies have linked the formation of PCs with elevated temperature in several plant species and C. elegans (Higgins et al. 2012; Bilgir et al. 2013). As the PCs observed in corollamau mutants always contained Corolla and Cona, we infer that their interaction was not disrupted. While 25 °C is the standard rearing temperature for both D. melanogaster and D. mauritiana, we hypothesized that the 74 amino acid changes in D. mauritiana Corolla could destabilize its interaction with D. melanogaster C(3)G and Cona in a temperature-dependent manner. Hydrophobic multivalent interactions have been shown to be important for phase-phase separation (Rog et al. 2017; Zhang et al. 2020; Kursel et al. 2021), and a lowering of temperature leads to a reduction of molecular motion and the stabilization of condensates (Jeon et al. 2025). Therefore, we asked if rearing corollamau mutants at 18 °C could rescue the early disassembly phenotype and the occurrence of PCs. Indeed, early SC disassembly was not observed in corollamau flies reared at 18 °C (Fig. 2b and Supplementary Fig. 9). Notably, PC formation was still observed in these germaria. Specifically, we observed PCs in 6/34 pro-oocytes at early pachytene, 8/13 at early-mid pachytene, and 2/2 at mid pachytene (Supplementary Fig. 8a). Interestingly, while the ratio of PCs with and without C(3)G was 39:61 (9/18 PCs for each type) for corollamau reared at 25 °C, for those reared at 18 °C we observe that only 3/19 PCs contain C(3)G while 16/19 PCs did not (Supplementary Fig. 8a). From this we conclude that the formation of PCs is not caused by early SC disassembly in corollamau mutants reared at 25 °C. We further observed fewer C(3)G-positive PCs in corollamau females reared at 18 °C. This may suggest that Corolla from D. mauritiana and Cona from D. melanogaster are prone to form PCs with each other, but as the SC is not disassembling early at 18 °C, C(3)G is not readily available to join the PCs.
As an alternative explanation for the formation of PCs in corollamau mutant germaria, we compared linear recognition motifs present on the Corolla proteins from D. melanogaster and D. mauritiana using the ELM resource for functional sites in proteins (Supplementary Table 1) (Kumar et al. 2024). Important to note here is that the corolla gene theoretically encodes for two isoforms of Corolla with only a three amino acid difference (Supplementary Fig. 10). Currently, it is unclear whether both isoforms are expressed, and functional studies should help clarify this. We also do not know which of the linear motifs mentioned below are functionally important for Corolla and the SC. Corolla isoforms A and B in D. melanogaster contain five and six functional sites, respectively, that are unique to the D. melanogaster protein. Candidates that could potentially create problems are the absence of a COP1 E3 ligase binding degron motif, a SUMO interaction site (both found on isoform A and B), as well as an APC/C destruction box motif (only on isoform B) on the Corolla protein from D. mauritiana (Supplementary Table 1). The lack of these motifs may affect transcriptional regulation and protein degradation. Importantly, the gene sina also codes for an E3 ligase. While the sina mutants studied in Hughes et al. are mutations in the gene body (Hughes et al. 2019), the lack of the required motif in the degradation target would likely also result in functional problems. In the Corolla protein of D. mauritiana there are two unique motifs, one of which is an additional motif for the deubiquitinase Usp7. While it was shown that the knockdown of Usp7 has detrimental effects on SC maintenance, recombination, and fertility (Lake et al. 2024), it is unclear what effects an additional Usp7 motif would have. To understand whether any of these differences in linear recognition motifs may lead to changes in how corolla is expressed in corollamau flies, we turned to quantitative polymerase chain reaction (qPCR). We did find a 19-fold overexpression of corolla in corollamau flies reared at 25 °C and a 12-fold overexpression in corollamau flies reared at 18 °C (Supplementary Fig. 11). The difference between corollamau flies reared at both temperatures was not statistically significant (Supplementary Fig. 11). Previously, it has been shown that the overexpression of corolla, achieved by the insertion of additional gene copy numbers, leads to the formation of PCs (Feijão et al. 2022). However, the morphology of the PCs reported by us compared to Feijão et al. is quite different. While our observed PCs were toroidal and hollow and started appearing in early pachytene, PCs reported by Feijão et al. are elongated filaments that begin forming as the SC disassembles in diplotene (Feijão et al. 2022). We cannot exclude that the overexpression of corollamau is linked to any of our observed phenotypes because the regulation of corolla gene expression has not been systematically studied. Further research will be needed to investigate the functional consequences of corolla overexpression at similar levels.
Chromosome segregation and recombination are significantly affected in corolla mau femalesGiven the effects on SC assembly, we next tested how the corolla gene replacement affected fertility. For this, we assessed the number of eclosed progeny from 10 females per genotype and rearing temperature in their first week. While flies at 18 °C overall had less progeny, no statistically significant differences were found when we compared corollamau flies to corolla+ flies reared at the same temperature (Supplementary Fig. 12). The reduction of progeny in flies reared at the lower temperature is expected due to slower oocyte development. After this, we assessed the effects on chromosome segregation and recombination. In corollamau flies, X chromosome missegregation increased significantly (Fig. 3a and Supplementary Fig. 13a). This effect was slightly less pronounced in flies reared at 18 °C (8.5% vs. 10.8% at 25 °C), but the difference was not statistically significant (Fig. 3a and Supplementary Fig. 13a). Importantly, at either temperature the rate of X chromosome missegregation was drastically lower than the 44.5% reported in corolla null mutant flies (Collins et al. 2014). 4th chromosome missegregation was significantly lower in corolla+ flies reared at 18 °C compared to 25 °C but, surprisingly, corollamau females did not show significant differences compared to corolla+ females (Fig. 3a and Supplementary Fig. 13a). This suggests that the proper segregation of the 4th chromosome in control females was improved at 18 °C compared to 25 °C. We would therefore have expected to see an increase in 4th chromosome missegregation in conjunction with the increase in X chromosome missegregation in corollamau mutants. The small and mostly heterochromatic 4th chromosome in D. melanogaster is, with very few exceptions, achiasmate and segregates via the distributive system (reviewed in (Scott Hawley and Theurkauf
Comments (0)