
Remember me
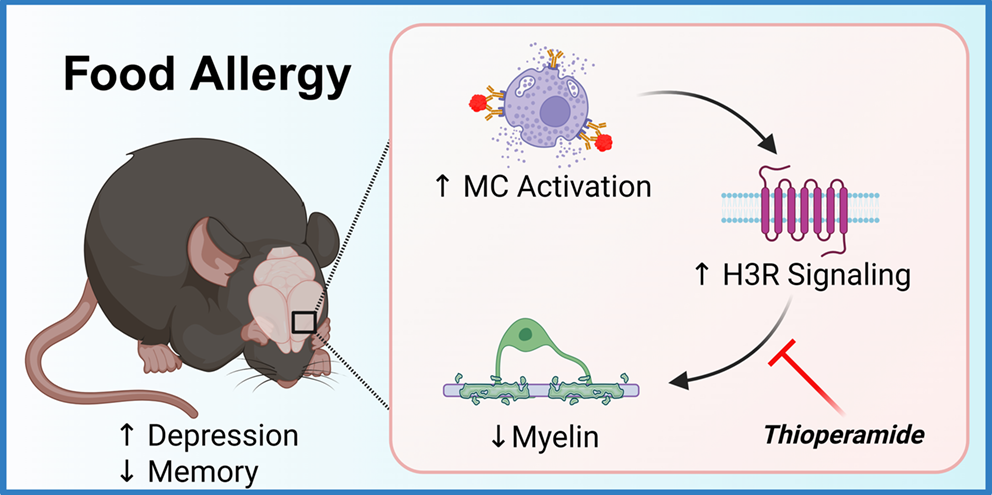

BLG-sensitized CMA mice did not show apparent physical symptoms indicative of food allergy-related reactions during the experimental period, as previously reported (Germundson and Nagamoto-Combs 2022). Both male and female BLG mice gained weight after sensitization (Week 5) and during allergen exposure and thioperamide (Thio) treatment (Week 7) at a similar rate to their sham counterparts (Fig. 1, b & c). Nevertheless, BLG-specific IgE (BLG-IgE) was significantly elevated in the sensitized mice overall, regardless of sex or thioperamide treatment (Fig. 2a; p < 0.001, 3-way ANOVA; also see Supplementary Fig. 2-1a). Multiple comparisons showed that the sensitization-induced increases in BLG-IgE were significant compared to their sex- and treatment-matched sham groups (Fig. 2b, significance indicated with numeric p-values, 3-way ANOVA). In addition to the sensitization status, sex also significantly affected BLG-IgE levels (Fig. 2a; p = 0.004, 3-way ANOVA), with male mice having overall greater allergen-specific antibody levels than females (Supplementary Fig. 2-1b; p = 0.0398, Student’s t-test). Thioperamide treatment did not affect the sensitization-induced BLG-IgE levels (Fig. 2, a & b).
Fig. 2
The levels of BLG-specific and total immunoglobulins and histamine (HA) and n-methylhistamine (NMHA) in the plasma. The circulating levels of BLG-IgE (a & b), BLG-IgG1 (c & d), total IgE (e & f), total IgG (g & h), total IgE/IgG ratio (i & j), and HA + NMHA (k & l) were quantified from the terminal plasma samples from male and female sham or BLG-sensitized mice treated either with the vehicle or thioperamide. The individual data values were calculated by subtracting the reference optical density (OD) values at 550 nm from the respective detection OD values at 450 nm. The IgE/IgG ratios were calculated by dividing the total IgE by the total IgG (l & j). All experimental groups were first analyzed by a 3-way ANOVA to identify significant main effects and interactions of sensitization, sex, and thioperamide treatment (treatment) as the independent variables, and the results were tabulated (a, c, e, g, i & k). The bold and italicized bold numbers indicate the F- and p-values of significant main effects and interactions, respectively (see Supplementary Figs. 2-1, 2-2, & 2-3 for post hoc analyses). Individual data values were also plotted to visualize the experimental outcomes (b, d, f, h, j & l). Each bar indicates the group average values ± SEM (male: n = 17–20 per group; female: n = 10–13 per group). Numbers between two bars indicate significant p-values from post hoc multiple comparisons. Sex-disaggregated analyses were also performed by 2-way ANOVA, and asterisks between two bars indicate significance (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001)
Likewise, BLG-IgG1 levels were similarly affected by sensitization and sex (Fig. 2c; sensitization: p < 0.001; sex: p = 0.007, 3-way ANOVA; also see Supplementary Fig. 2-1, c & d). Furthermore, there was a significant interaction between sex and sensitization (p = 0.004). Indeed, BLG-IgG1 levels were significantly higher in sensitized females than sensitized males (Fig. 2d; green vs. orange filled bars, p = 0.0008, 3-way ANOVA). The interaction of sex and sensitization was further tested among the male and female subgroups (Supplementary Fig. 2-1e). While no difference in the mean values for BLG-IgG1 was observed between male and female sham groups (male sham = 0.22 ± 0.03 vs. female sham = 0.22 ± 0.03; p = 0.9761, 2-way ANOVA), the difference was significant for BLG-sensitized males and females (male BLG = 0.45 ± 0.05 vs. female BLG = 0.69 ± 0.05; p = 0.0002, 2-way ANOVA), even though the level was also elevated in sensitized males compared to their respective sham mice (Fig. 2d; open and filled green bars, p = 0.0210, 3-way ANOVA, ****p < 0.0001, 2-way ANOVA). To note, thioperamide seemed to elevate the overall levels of BLG-IgG1 slightly (Fig. 2c; p = 0.049, 3-way ANOVA), although the antagonist did not alter the sensitization-induced BLG-IgG1 levels within each sex (Fig. 2d). These results indicated that, while thioperamide treatment during the allergen exposure did not influence the production of BLG-specific IgE and IgG1 in both sexes, allergen-specific IgG1 might be produced more preferentially in female mice than in males with allergen exposure.
We also measured total IgE and IgG from terminal plasma samples to assess whether BLG sensitization and thioperamide treatment affected overall immunoglobulin production (Fig. 2, e-j). In a sex-aggregated analysis, a significant effect of BLG sensitization on total IgE levels was found (Fig. 2e; p = 0.006, 3-way ANOVA; also see Supplementary Fig. 2-2a). Within males, total IgE in vehicle-treated BLG mice was significantly greater than in sham mice (Fig. 2f; open vs. filled green bars: **p = 0.0063, 2-way ANOVA), but the difference between vehicle-treated female sham and BLG mice was not significant (Fig. 2f; open vs. filled orange bars). Similarly, no differences were observed among the thioperamide-treated groups. We did not find any notable differences among the groups for total IgG1 levels (Fig. 2, g & h). Additionally, while there appeared to be a main effect of sensitization on the overall IgE/IgG ratio (Fig. 2i; p = 0.044, 3-way ANOVA), post hoc tests showed that the difference between sham and BLG-sensitized animals did not reach a statistical significance (Supplementary Fig. 2-2b; p = 0.0738, Student’s t-test).
The levels of HA and its metabolite N-methylhistamine (NMHA) were also quantified in the plasma samples isolated at the time of euthanasia as an additional indicator of hypersensitivity response (Fig. 2, k & l). The sensitization status of the animals significantly affected the levels of HA + NMHA (Fig. 2k; p < 0.001, 3-way ANOVA; also see Supplementary Fig. 2-3a), increasing the plasma levels of HA + NMHA in sensitized mice of both sexes (Fig. 2l). Sex (p < 0.001, 3-way ANOVA) and thioperamide treatment (p = 0.032, 3-way ANOVA) were also independent variables that significantly affected the levels of the analytes (Fig. 2k). However, post hoc comparisons between male and female groups (Supplementary Fig. 2-3b) or between vehicle and thioperamide treatment groups (Supplementary Fig. 2-3c), indicated that the effect of sex (p = 0.0009), but not treatment (p = 0.1019), was significant, suggesting that other independent variables were confounding factors. Although a 3-way ANOVA multiple comparisons indicate that the difference in HA + NMHA levels between sensitized male and female mice was significant when compared within the thioperamide-treated group (Fig. 2l; purple vs. pink filled bars, p = 0.0115, 3-way ANOVA), the interaction between the sex and treatment was not significant (Fig. 2k). These results validated that H3R inhibition did not diminish peripheral HA release in sensitized mice during whey-protein-diet consumption.
Effects of Thioperamide Treatment On CMA-associated Depression-like Behavior and Recognition Memory in Male Mice Sensorimotor functionAt the start of Week 7, all mice underwent a series of behavioral tests over 4 consecutive days. Mice were subjected to the grip strength test (Fig. 3, a & b) and rotor-rod test (Fig. 3, c & d) to determine whether BLG sensitization and thioperamide treatment altered motor function and coordination, respectively. No significant overall differences were detected due to sensitization (Fig. 3, a-d; 3-way ANOVA) or when BLG-sensitized mice were compared to their treatment-matched sham controls within the same sex (Fig. 3, b & d; 2-way ANOVA). However, thioperamide-treated male BLG mice showed slightly greater grip strength compared to sex-matched vehicle-treated BLG mice (Fig. 3b; *p = 0.0136, 2-way ANOVA), which was not observed in female mice (Supplementary Fig. 3-1, a & b). For the rotor-rod test, there was a small but significant main effect of sex on the latency to fall (Fig. 3c; p = 0.035). A post hoc analysis validated that the differences between the sexes were significant (Supplementary Fig. 3-1c, p = 0.0366, Student’s t-test), indicating that females generally stayed on the equipment slightly longer than males.
Fig. 3
Sensorimotor function assessments with the grip strength and rotor-rod tests and locomotion and anxiety-like behavior assessments with the open field test. During Week 7, male and female sham and BLG mice treated with vehicle or thioperamide were subjected to the grip strength test (a & b) and rotor-rod test (c & d) to measure their limb strengths and sensorimotor function, respectively. Grip strength (a & b) and rotor rod tests (c & d) were conducted as 3 separate sequential trials, and the average of all trials was presented as the final result for each group. The open field test was also used to test the animals’ locomotion and anxiety-like behaviors (e-p). During the open field test, mice were allowed to move freely for 10 min to measure their general activity (e-l) and exploratory/anxiety-like behaviors (m-p). The results from the first 5 min of the open field test were presented as the final results. All experimental groups were first analyzed by a 3-way ANOVA to identify significant main effects and interactions of sensitization, sex, and thioperamide treatment (treatment) as the independent variables, and the results were tabulated (a, c, e, g, i, k, m & o). The bold numbers indicate the F- and p-values for sex as a significant main effect (see Supplementary Fig. 3-1, 3-2, & 3-3 for post hoc analyses). Individual data values were also plotted to visualize the experimental outcomes (b, d, f, h, j, l, n & p). Each bar indicates the group average values ± SEM (male: n = 17-20 per group; female: n = 10-13 per group). Sex-disaggregated analyses were also performed by 2-way ANOVA, and asterisks between two bars indicate significance (*p < 0.05)
Exploratory behaviorTo assess overall activities, including mobility, exploration, and location preference, the animals were also subjected to the open field test. A set of sex-aggregated analyses revealed significant effects of sex for parameters concerning immobility (Fig. 3, e & f; time immobile: p < 0.001; Fig. 3, g & h; number of immobile episodes: p < 0.001, 3-way ANOVA). Post hoc analyses comparing the sexes indicated that female mice were generally less mobile than male mice during the test (Supplementary Fig. 3-2 , a & b). Furthermore, significant interactions between sex and treatment were observed for these measures (Fig. 3 e & g; 3-way ANOVA), with thioperamide promoting immobility in sensitized females (Supplementary Fig. 3-3, a & b). In addition, significant interactions between sensitization and treatment were observed both for the total distance traveled (Fig. 3i; p = 0.025) and for the average speed (Fig. 3k; p = 0.023), with thioperamide reducing the values of these parameters in sham mice and the difference between sham and sensitized mice, regardless of sex (Supplementary Fig. 3-3, c & d).
Additional sex-disaggregated analyses confirmed that male BLG mice traveled shorter distances (Fig. 3j, open vs. filled green bars; **p = 0.0082, 2-way ANOVA) and moved at lower speeds (Fig. 3l, open vs. filled green bars; **p = 0.0073, 2-way ANOVA), but these sensitization-associated differences in distance and speed were not significant in females (Fig. 3, j & l, open vs. filled orange bars). On the other hand, the measures reflective of anxiety-like behavior with the open field test, such as time spent in the center zone (Fig. 3, m & n) and the number of center zone entries (Fig. 3, o & p), did not differ between sham and BLG mice of either sex. Interestingly, thioperamide treatment did not further reduce these activities reflective of mobility and exploratory behaviors in BLG mice.
Thus, while BLG sensitization did not impair sensorimotor function in either sex, it resulted in significant changes in some aspects of male behavior. Furthermore, the effect of thioperamide on behavior was differentially influenced by sex and the sensitization status of mice, depending on the specific behavioral parameters being assessed.
Depression-like behaviorDuring the behavioral testing period, mice were also evaluated for their depression-like behavior using the tail suspension test (Fig. 4, a-d). BLG sensitization did not significantly influence the overall test outcomes of the cohort, but sex and thioperamide treatment significantly affected the latency to time immobile (Fig. 4c; sex: p = 0.001; treatment: p = 0.004, 3-way ANOVA). Moreover, there was a significant interaction between sex and treatment that affected the latency (p = 0.028, 3-way ANOVA). A post hoc analysis of the 3-way ANOVA indicated that, among the thioperamide-treated groups, the latency to immobility was greater in female mice than in males during the tail-suspension test, regardless of their sensitization status (Supplementary Fig. 4-2e; p < 0.0001, 2-way ANOVA).
Fig. 4
Depression-like behavior analysis with the tail suspension test and cognitive function assessments with cross maze and novel object recognition tests. During Week 7, male and female sham and BLG mice treated with vehicle or thioperamide were subjected to the tail suspension test for 6 min to assess depression-like behaviors, including the time immobile (a & b) and the latency to their first immobile episode (c & d). Additionally, mice were subjected to the cross maze test (e & f) and the novel object recognition test (g & h) to assess their short-term spatial and object-recognition memory, respectively. For the analysis of novel object recognition (g & h), the recording from the first 5 min of the 10-min testing period was used for the final results (see Supplementary Fig. 4-1 ). For each of the parameters measured, all experimental groups were first analyzed by a 3-way ANOVA to identify significant main effects and interactions of sensitization, sex, and thioperamide treatment (treatment) as the independent variables, and the results were tabulated (a, c, e, g). The bold numbers and bold italicized numbers indicate the F- and p-values of significant main effects and interactions, respectively (see Supplementary Figs. 4-2 and 4-3 for post hoc analyses). Individual data values were also plotted to visualize the experimental outcomes (b, d, f, h). Each bar indicates the group average values ± SEM (male: n = 17–20 per group; female: n = 10–13 per group). Numbers between two bars indicate significant p-values from post hoc multiple comparisons. Sex-disaggregated analyses were also performed by 2-way ANOVA, and asterisks between two bars indicate significance (*p < 0.05, **p < 0.01)
When sham and BLG mice were further compared within their respective sex, we observed that vehicle-treated male BLG mice spent more time immobile during the test than their sham counterparts (Fig. 4b; open vs. filled green bars, *p = 0.0384, 2-way ANOVA), as described in our prior study (Germundson and Nagamoto-Combs 2022). The male BLG group also showed a trend of becoming immobile earlier, although this measurement was not statistically different from the sham group (Fig. 4d; open vs. filled green bars, p = 0.2281; 2-way ANOVA). The thioperamide treatment prevented the development of depression-like behavior in male BLG mice, showing a significant reduction in their time immobile compared to their respective vehicle-treated sensitized mice (Fig. 4b; green vs. purple filled bars, **p = 0.0050, 2-way ANOVA). However, no significant differences concerning their time immobile or latency to time immobile were observed between treatment-matched female sham and BLG mice (Fig. 4, b & d). Our results showed that male and female sensitized mice displayed unique changes in behavior with the tail suspension test, and that H3R antagonism attenuated this behavioral change at least in male BLG mice.
Spatial and novel object recognition memoryIn our earlier study, we reported a significant decrease in myelin density in the cerebral cortex of male BLG-sensitized mice (Germundson and Nagamoto-Combs 2022). As cortical demyelination is associated with declines in memory and cognition in multiple sclerosis (Kutzelnigg et al. 2005; Lucchinetti et al. 2011), we tested short-term spatial memory and object-recognition memory using the cross maze test and novel object recognition test, respectively. When measuring the percentage of successful alternations with the cross maze as an indication of short-term spatial memory, sex was not a significant main effect (Fig. 4e; 3-way ANOVA). However, post hoc pairwise multiple comparisons within the thioperamide-treated groups indicated that female sham mice scored slightly but significantly lower than male sham mice (Fig. 4f; p = 0.0458, 3-way ANOVA). When males and females were further analyzed within respective sexes, however, no significant differences were observed among any of the male or female treatment groups (Fig. 4f).
With the novel object recognition test, a significant main effect of sensitization was evident on the discrimination index (Fig. 4g; p = 0.047, 3-way ANOVA). However, a direct comparison between overall sex-aggregated sham and BLG-sensitized groups indicated that the difference between the groups did not reach statistical significance (Supplementary Fig. 4-3, p = 0.1651, Student’s t-test), suggesting that the significant main effect of sensitization was influenced by the combination of sex and treatment factors. In support of this interpretation, post hoc multiple comparisons indicated that the differences in discrimination indices were significant between the vehicle-treated and thioperamide-treated male BLG groups (Fig. 4h; green vs. purple filled bars, p = 0.0231, 3-way ANOVA) as well as between thioperamide-treated male and female sham groups (Fig. 4h; purple vs. pink open bars, p = 0.0462, 3-way ANOVA). The F-value for the interactions among sensitization, sex, and treatment was 3.41, although it did not reach statistical significance (Fig. 4g; p = 0.0673, 3-way ANOVA),
In addition to validating the above-mentioned findings, a sex-disaggregated analysis further indicated that the discrimination index of vehicle-treated sensitized male mice was significantly lower than that of treatment-matched sham male mice (Fig. 4h; *p = 0.0379, 2-way ANOVA). The difference between the vehicle- and thioperamide-treated male BLG mice was also validated with this analysis (Fig. 4h; green vs. purple filled bars, *p = 0.0159, 2-way ANOVA), suggesting that H3R inhibition showed a beneficial effect on their short-term memory by improving object recognition. Female BLG mice showed only a decreasing trend in the discrimination index in a sex-disaggregated analysis (Fig. 4h; p = 0.1837, 2-way ANOVA), and thioperamide treatment did not appreciably improve this trend (p = 0.0924, 2-way ANOVA). Despite the tendency of sham females to perform better than their treatment-matched male counterparts (Fig. 4h), only the difference between thioperamide-treated sham males and sham females was significant (Fig. 4h; p = 0.0462; 3-way ANOVA).
Taken together, these findings indicated that thioperamide treatment improved some aspects of the CMA-associated behavioral changes and cognitive declines, and the effect of H3R inhibition on behavioral changes was more demonstrable in male mice.
Effects of Thioperamide Treatment On the Activation or IgE Coupling of Intracranial Mast Cells in BLG-Sensitized MicePreviously, we showed that mast cells, metachromatically stained with acidic toluidine blue, were prevalent throughout the dura mater of the meninges regardless of their sensitization status, with more mast cells degranulated in BLG mice (Germundson and Nagamoto-Combs 2022). We also demonstrated that numerous FcεRI-immunoreactive cells were present in both sham and BLG-sensitized animals, but the number of IgE-immunopositive cells was appreciable only in sensitized mice. To confirm that thioperamide attenuated the CMA-associated behavioral changes via its inhibitory action on H3R and not via unexpected effects on intracranial mast cell properties, we investigated whether thioperamide influenced the degranulation status of dural mast cells and their coupling with IgE. For this purpose, we co-stained dural mast cells for CD117 (c-kit) and IgE and examined their morphology to assess whether IgE-coupled mast cells had been degranulated.
Total mast cells in the duraIn our initial observation, CD117-immunopositive mast cells were found throughout the dura mater regardless of the sensitization condition or sex (Fig. 5, a & b). However, when mast cells were quantified, BLG sensitization was found to slightly but significantly influence the overall number of dural mast cells (Fig. 5c; p = 0.042, 3-way ANOVA). Furthermore, an interaction between sensitization and sex (p = 0.029) was identified, and a post hoc test revealed that the elevation in the total number of mast cells was specific to BLG males compared to sham males (Supplementary Fig. 5-1a; p = 0.0028, 2-way ANOVA), and the difference between male and female sensitized mice was significant (Supplementary Fig. 5-1a; male BLG = 65 ± 3 vs. female BLG = 55 ± 3; p = 0.0294, 2-way ANOVA), although no significant change was noted between sexes of sham mice (male sham = 52 ± 3 vs. female sham = 56 ± 3; p = 0.3233, 2-way ANOVA). When compared among groups within each sex, only the difference between thioperamide-treated sham and BLG male mice was significant, with 25% more mast cells in the latter group (Fig. 5d; *p = 0.0380, 2-way ANOVA).
Fig. 5
Immunofluorescence staining and quantification of IgE-coupled intracranial mast cells. Terminal dura mater tissues from male (a) and female (b) sham and BLG mice treated with vehicle or thioperamide were immunofluorescently triple-stained for CD117 (red), IgE (green), and DAPI (blue). Representative photomicrographs of mast cells (MC) are shown. The number of total CD117+ mast cells (c-h) and IgE+ mast cells (i-n) was quantified by counting the stained cells in 20 distinct microscope fields at 40× magnification, and % IgE+ mast cells was calculated. Scale bar = 20 μm. For each mast cell phenotype, all experimental groups were first analyzed by a 3-way ANOVA to identify significant main effects and interactions of sensitization, sex, and thioperamide treatment (treatment) as the independent variables, and the results were tabulated (c, e, g, i, k & m). The bold and italicized bold numbers indicate the F- and p-values of significant main effects and interactions, respectively (see Supplementary Figs. 5-1 and 5-2 for post hoc analyses). Individual data values were also plotted to visualize the experimental outcomes (b, d, f, h, j, l & n). Each bar indicates the group average values ± SEM (n = 9). Numbers between two bars indicate significant p-values from post hoc multiple comparisons. Sex-disaggregated analyses were also performed by 2-way ANOVA, and asterisks between two bars indicate significance (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001). See Supplementary Fig. 5-3 for correlation plots between the levels of BLG-IgE and the number/percentage of mast cells of distinct morphology
Based on these findings, we further quantified granulated or degranulated mast cells based on their morphology. No significant effects were found on the number of granulated mast cells in a sex-aggregated analysis (Fig. 5e). When sexes were compared independently, BLG sensitization significantly decreased the number of granulated mast cells across all groups, except the thioperamide-treated female group having a higher number of granulated mast cells than their sex-matched vehicle controls (Fig. 5f; orange vs. pink filled bars, *p = 0.0166, 2-way ANOVA).
In contrast to granulated mast cells, the number of degranulated mast cells in the dura was strongly affected by sensitization status (Fig. 5g; p < 0.001, 3-way ANOVA) as we observed for the total mast cell numbers (see Fig. 5c). An interaction between sex and sensitization status also significantly influenced the number of degranulated mast cells (Fig. 5g; p < 0.001, 3-way ANOVA). Post hoc multiple comparisons indicated that BLG sensitization markedly increased the number of mast cells with the activated morphology in male sensitized mice compared to sex-matched sham and to female BLG mice (Supplementary Fig. 5-1b; male sham = 14 ± 2 vs. male BLG = 28 ± 2, p < 0.0001; male BLG = 28 ± 2 vs. female BLG = 20 ± 2, p = 0.0085, 2-way ANOVA). The difference between male and female sham groups was not significant (male sham = 14 ± 2 vs. female sham = 19 ± 1; p = 0.1255, 2-way ANOVA).
In a sex-disaggregated analysis, the sensitization-associated increases in the number of degranulated mast cells were apparent in the vehicle-treated mice of both sexes (Fig. 5h; male: sham vs. BLG, ***p = 0.0002; female: sham vs. BLG, *p = 0.0290, 2-way ANOVA). Although the 3-way ANOVA did not detect significant main effect or interactions of treatment, direct comparisons of vehicle-treated and thioperamide-treated groups within the same sex indicated that the H3R antagonist affected degranulated mast cell number in female mice, decreasing the sensitization-associated changes (Fig. 5h; orange vs. pink filled bars, **p = 0.0084, 2-way ANOVA), having an opposite effect as it did for granulated cells (see Fig. 5f). Thioperamide-treated sensitized male mice still showed significant increases in degranulated mast cell number compared to the treatment-matched sham mice (Fig. 5h; open and filled purple bars, *p = 0.0166, 2-way ANOVA).
IgE-immunoreactive mast cells in the duraWhen dural mast cells were further characterized by their double-immunoreactivity to both CD117 and IgE, the overall percentage of IgE-positive (IgE+) mast cells was impacted not only by sensitization (Fig. 5i; p < 0.001, 3-way ANOVA) but also by sex (p = 0.004, 3-way ANOVA), with BLG sensitization and male sex independently increasing the overall percentage of IgE+ mast cells (Supplementary Fig. 5-2, a & b, respectively). In addition, significant interactions between sensitization and sex (Fig. 5i; p = 0.005) and sensitization and treatment (p = 0.016) were identified. When the interaction between sex and sensitization was further analyzed, the percentage of total IgE+ mast cells in BLG-sensitized males was significantly greater than the female BLG group (Supplementary Fig. 5-2c; male BLG = 66 ± 5% vs. female BLG = 41 ± 4%, p = 0.0008, 2-way ANOVA), indicating a strong sex-specific effect in terms of the sensitization and treatment interaction. In addition, post hoc multiple comparisons indicated that the percentage of IgE-coupled mast cells was elevated in vehicle-treated sensitized mice of both sexes (Fig. 5j; male: open vs. closed green bars, p = 0.0001; female: open vs. filled orange bars, p = 0.0229, 3-way ANOVA). While thioperamide did not decrease the sensitization-associated increase in the IgE+ mast cells in males (Fig. 5j; open vs. filled purple bars, p = 0.0070, 3-way ANOVA), the H3R antagonist decreased this population of mast cells in female BLG mice compared to the treatment-matched sham (Fig. 5j, open vs. filled pink bars) or vehicle-treated female counterparts (Fig. 5j, orange vs. pink filled bars, p = 0.0174, 3-way ANOVA), suggesting that the H3R antagonist reduces the number of allergen-responsive mast cells in female dura.
An additional morphological assessment of double immunopositive cells showed that there was no change in the percentage of IgE-coupled granulated mast cells (Fig. 5, k & l). However, large percentages of IgE-positive mast cells were degranulated in BLG-sensitized mice (Fig. 5, m & n). Similarly to our findings with the overall IgE-coupled mast cells, IgE-associated degranulated mast cells were affected independently by sensitization (Fig. 5m; p < 0.001, 3-way ANOVA) and sex (p = 0.002, 3-way ANOVA). Post hoc analyses of these factors indicated that BLG sensitization and male sex significantly increased the percentage of IgE+ mast cells (Supplementary Fig. 5-2, e & f, respectively). We also found strong interactions between sensitization and sex (p < 0.001, 3-way ANOVA), as well as sensitization and thioperamide treatment (p = 0.002, 3-way ANOVA). Post hoc tests validated that the BLG-sensitized male group showed an elevated percentage of degranulated IgE+ mast cells compared to BLG-sensitized females (Supplementary Fig. 5-2g; male BLG = 32 ± 3% vs. female BLG = 16 ± 2%, p < 0.0001, 2-way ANOVA). The percentage between male and female sham groups was comparable (male sham = 10 ± 1% vs. female sham = 13 ± 2%, p =0.6998, 2-way ANOVA). Additional analyses of the interaction between sensitization and treatment indicated that sensitization-induced increases in IgE+ mast cell percentage were only significant in the vehicle-treated mice and not in the thioperamide-treated groups (Supplementary Fig. 5d-h; p = 0.0001, 2-way ANOVA). Multiple comparisons revealed that the effect of thioperamide to reduce the sensitization-induced increases in the percent of degranulated IgE+ mast cells was largely observed in female BLG groups and not in male mice (Fig. 5n; green vs. purple filled bars, p < 0.0001, 3-way ANOVA), suggesting that sex contributes as a confounding factor to some extent.
While the sex-aggregated analysis indicated that the H3R antagonist slightly increased the percent of degranulated IgE+ mast cells in sham females compared to the vehicle-treated counterpart (Fig. 5n; orange vs. pink open bars, p = 0.0137, 3-way ANOVA), the overall the difference between the two groups was no longer significant when analyzed by 2-way ANOVA within female groups. Nevertheless, thioperamide treatment significantly reduced the percentage of degranulated IgE+ mast cells in BLG-sensitized females compared to the vehicle-treated female BLG mice (Fig. 5n, orange and pink filled bars, **p = 0.0084, 2-way ANOVA). These findings indicate that IgE-coupled mast cells increase in the dura mater of sensitized mice regardless of sex, but their number may be uniquely regulated, with thioperamide having a notable influence in females.
Sex differences in the effect of sensitization became apparent when the relationships of the serum levels of BLG-specific IgE with the numbers of total mast cells, degranulated mast cells, and IgE-coupled dural mast cells were examined. We found significant correlations between BLG-IgE and these mast cell measures in males (Supplementary Fig. 5-3, a-f), but such correlations were not found in female mice (Supplementary Fig. 5-3, g-l). These observations indicated that the increased number and activation of mast cells were more likely associated with BLG sensitization in males than in females. Importantly, thioperamide did not alter sensitization-induced changes in CD117-positive mast cells in males, suggesting that the attenuation of behavioral changes observed with thioperamide treatment was not due to its influence on the activation or IgE coupling of dural mast cells.
Region-Specific Effects Of BLG Sensitization and Thioperamide on HA, N-methylhistamine, and Histamine N-methyltransferase Levels in the BrainBecause BLG sensitization significantly elevated the number of degranulated dural mast cells (Fig. 5, h & n), we predicted that excessive HA had been released into the intracranial space. Thus, we next quantified the HA levels in different brain regions. Using protein lysates from the frontal cortex (fCTX), hippocampus (HPC), and thalamus (THAL), we performed a competitive ELISA that detected both HA and its metabolite, N-methylhistamine (NMHA).
When HA + NMHA levels in the three brain regions were examined by 3-way ANOVA in sex-aggregated analyses, no overall significant main effect of sex was found in the frontal cortex with 3-way ANOVA (Fig. 6a). However, there was a significant interaction between sex and thioperamide treatment in the frontal cortex (p = 0.018, 3-way ANOVA). Multiple comparisons showed that the levels of HA + NMHA were markedly greater in sham females than sham males, regardless of thioperamide treatment (Fig. 6b; green vs. orange open bars, p = 0.0151; purple vs. pink open bars, p = 0.0097; 3-way ANOVA). Post hoc analyses on the interaction between sex and treatment indicated that the overall amounts of HA + NMHA in the frontal cortex were greater in female groups than in male groups (Supplementary Fig. 6-1a; male thio vs. female thio, p = 0.0001, 2-way ANOVA). We also found that, in males, the lower analyte levels in mice were significantly elevated after thioperamide treatment (Supplementary Fig. 6-1a; male vehicle vs. thioperamide, p = 0.0235, 2-way ANOVA). In a sex-disaggregated analysis, we found that the analyte levels were significantly elevated in BLG male mice compared to their sham counterpart (Fig. 6b, open vs. filled green bars, *p = 0.0469, 2-way ANOVA), and thioperamide treatment increased HA + NMHA levels in the frontal cortex of sham males (Fig.
Comments (0)