
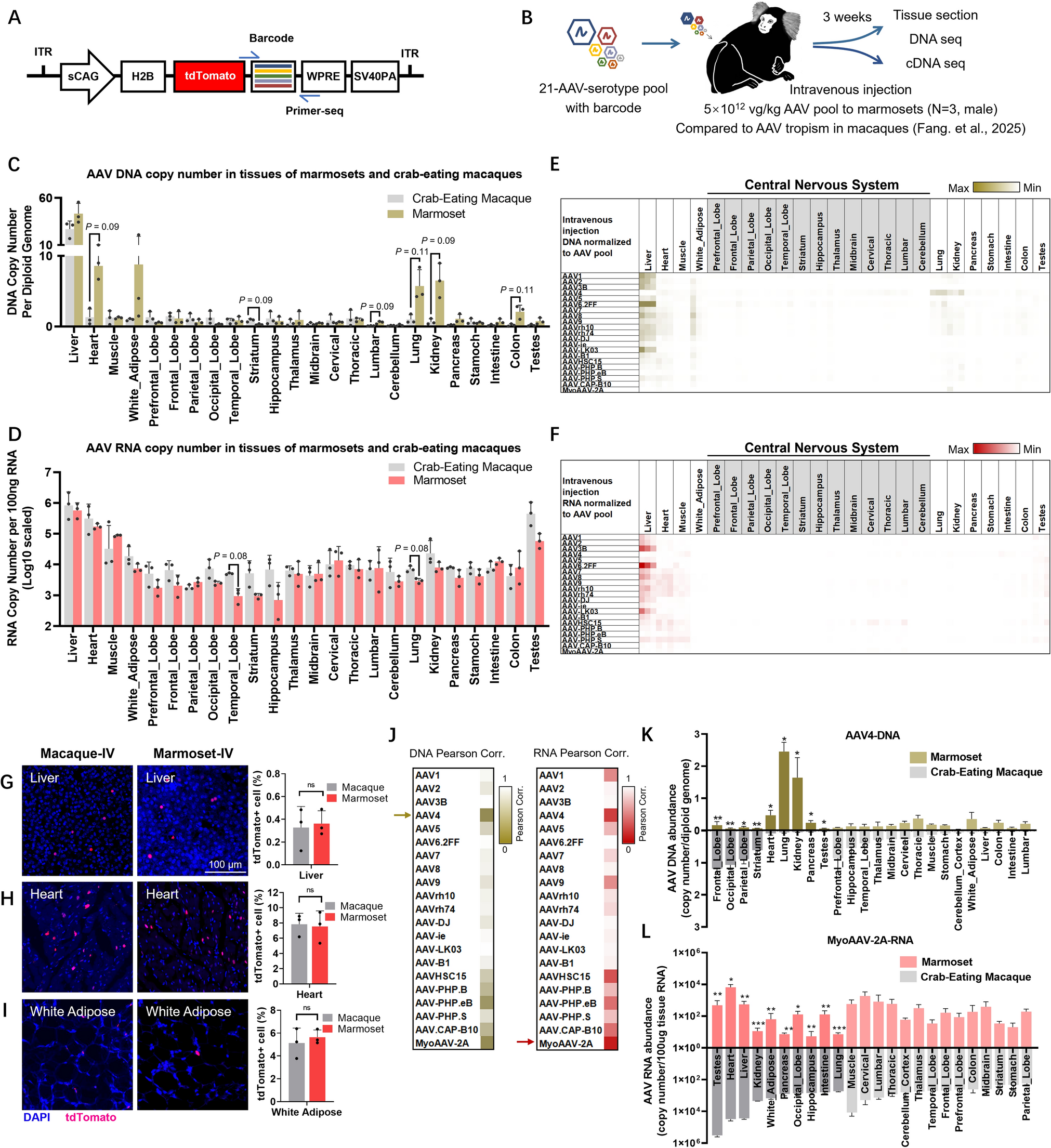
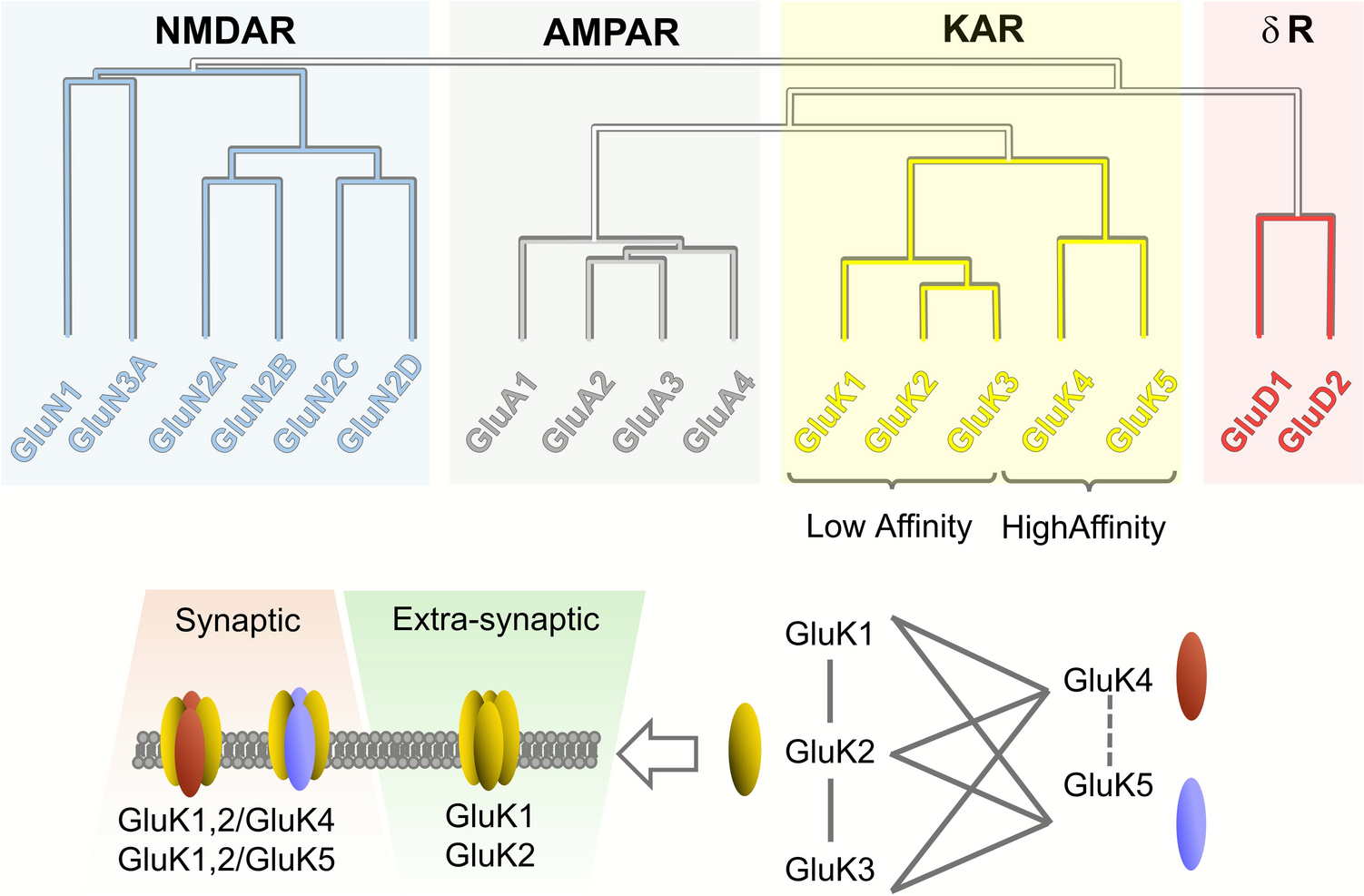
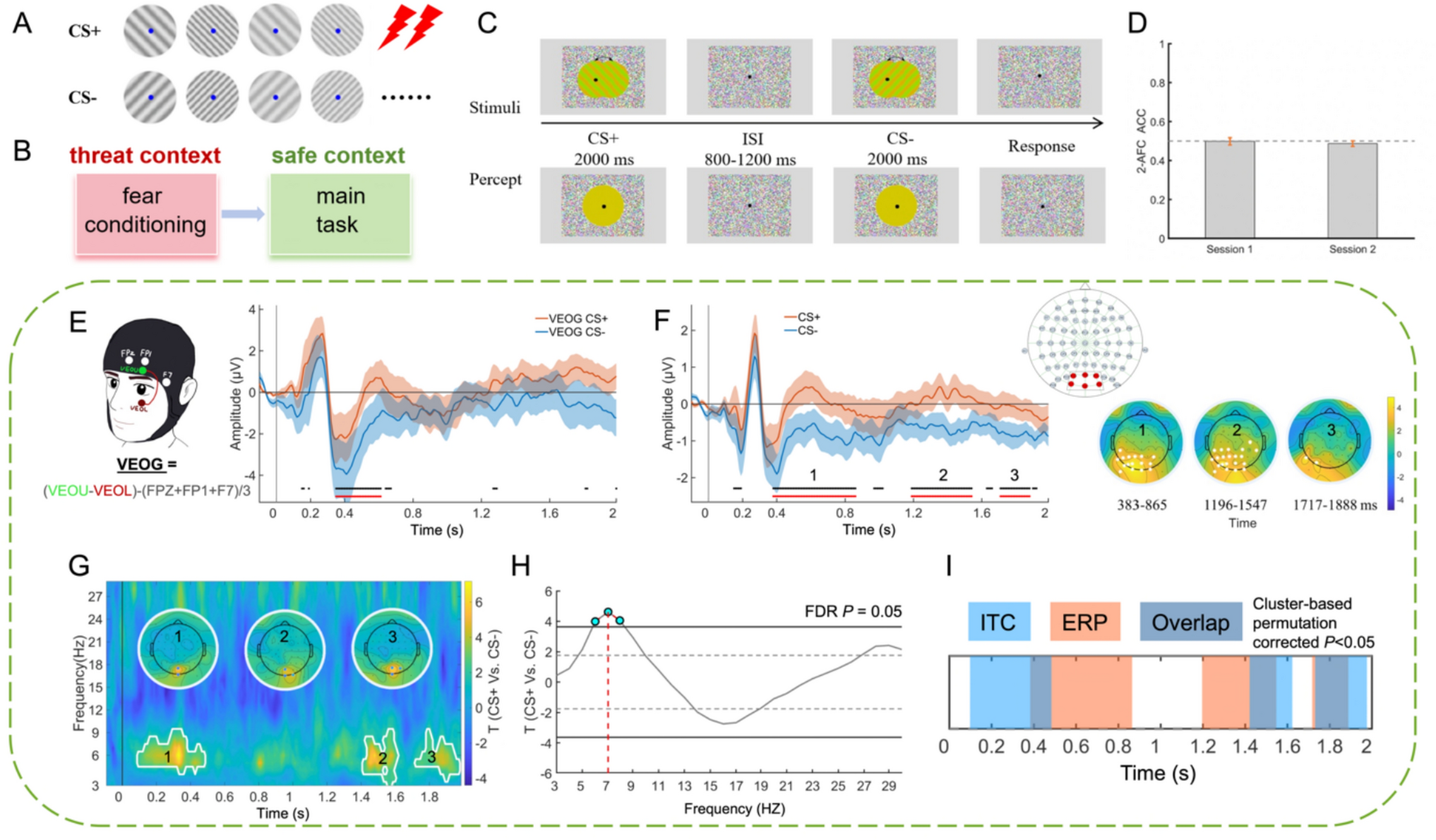
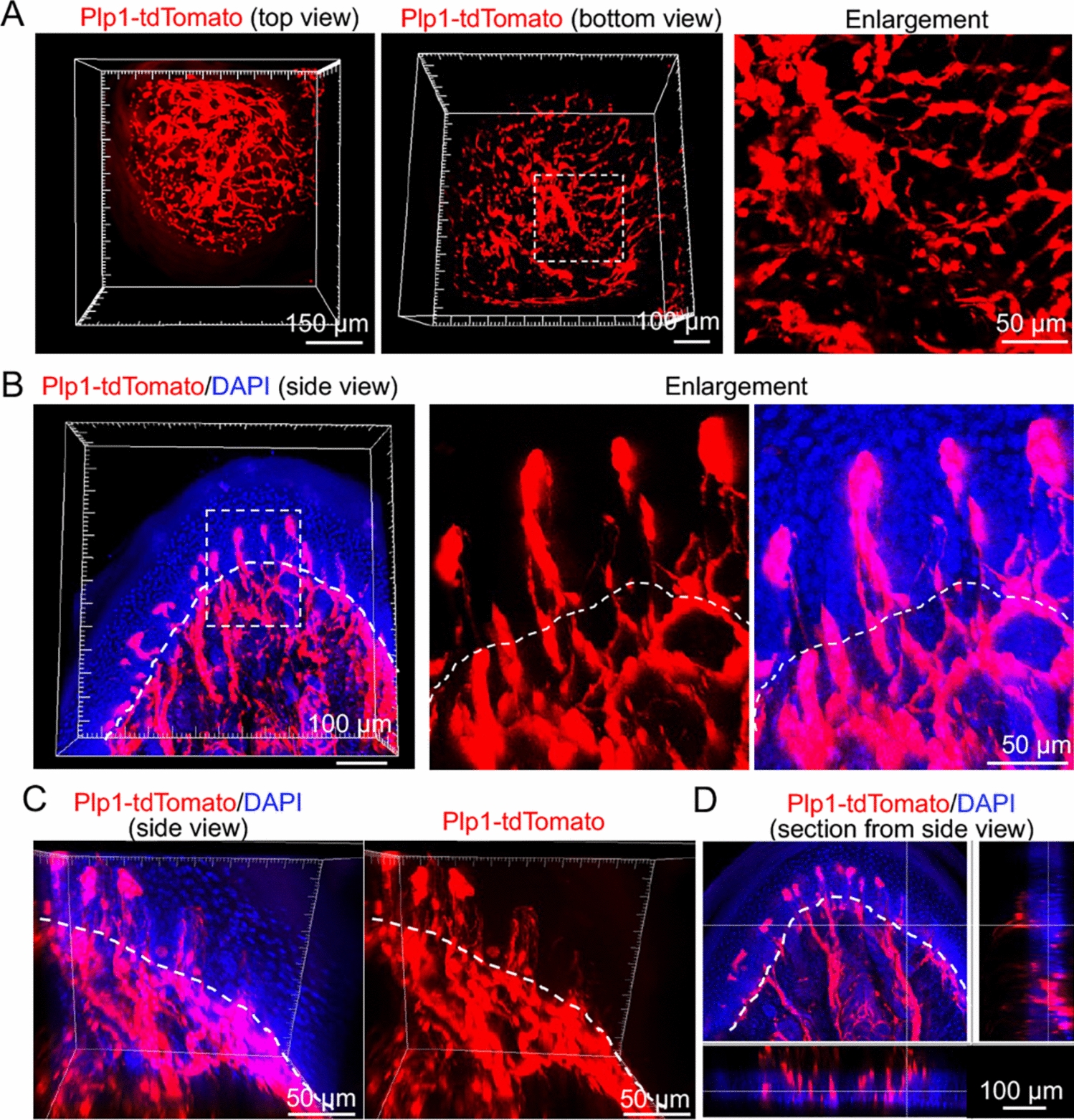
Remember me
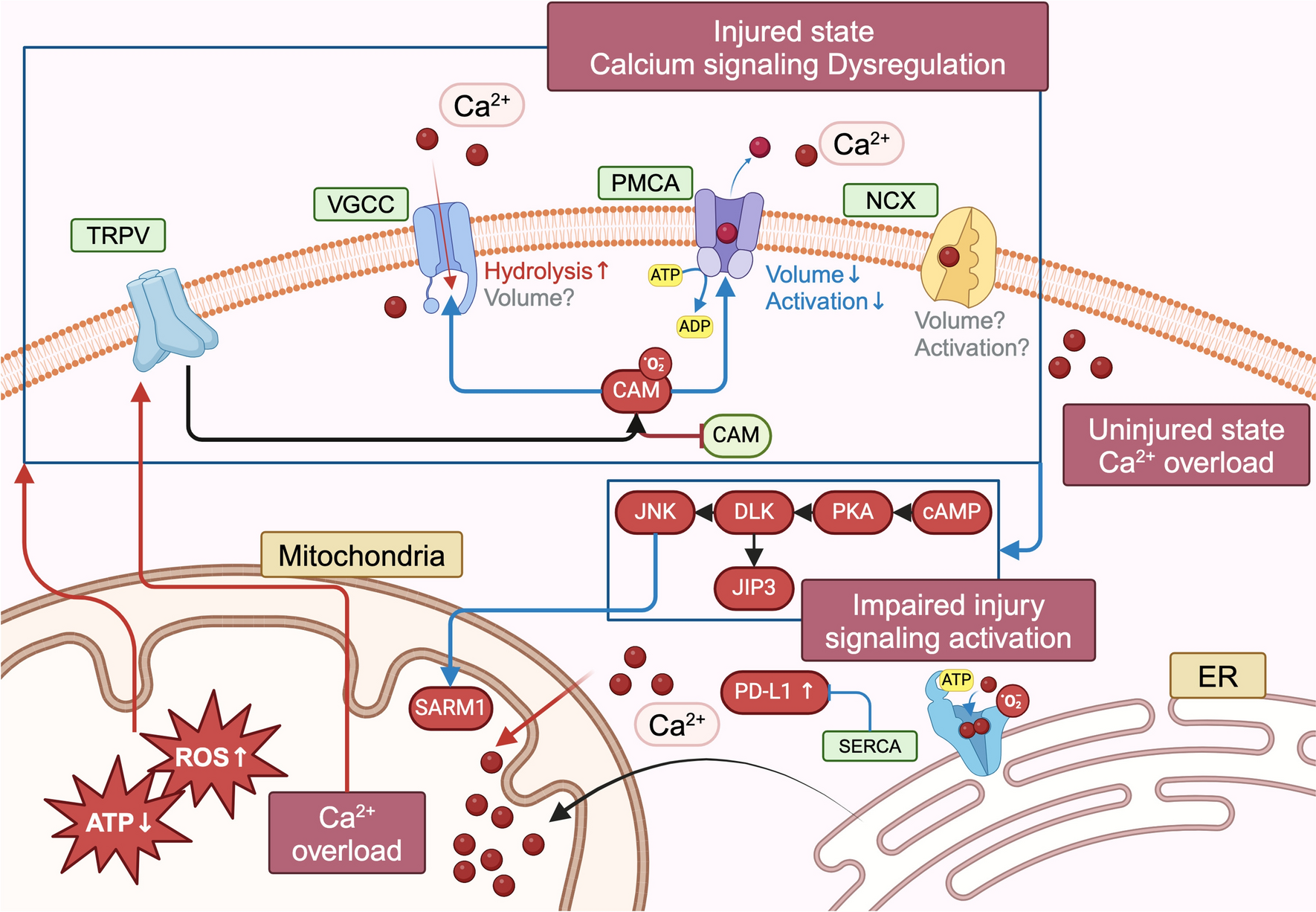

In the present study, we recorded 205 well-isolated single units from the IC of two awake marmosets. Among them, 164 neurons exhibited significant auditory responses to the acoustic flutter stimuli. To study temporal processing at the flutter range (10–45 pps), we delivered acoustic pulse trains at repetition rate in the range of 4–48 pps that varied linearly in 4-pps step (Fig. 1A). As shown in Figure 1B, there are four types of IC neural response to acoustic flutter based on their firing rate and temporal periodicity: positive-monotonic synchronized response (syn+), negative-monotonic synchronized response (syn-), positive-monotonic non-synchronized response (nonsyn+), and negative-monotonic non-synchronized response (nonsyn−), which is consistent with previous studies in the IC examined with larger time scale of repetition rate (10–500 pps) [12] and the AC examined at the flutter range [2]. In brief, both syn+ and syn- neurons exhibited precise click-synchronized spiking activity with a vector strength (VS) > 0.1 and Rayleigh statistics (RS) > 13.8 at most or all repetition rates in the flutter range (Fig. S1A–B). In contrast, nonsyn+ and nonsyn- neurons did not show stimulus-synchronized spiking activity (Fig. S1C–D, RS < 13.8). Thus, stimulus-synchronized neurons (syn, including syn+ and syn–) could be clearly distinguished from non-synchronized neurons (nonsyn, including nonsyn+ and nonsyn–) by RS (Fig. 1C and Fig. S1). In addition, the number of syn neurons (n = 96) is more than the number of nonsyn neurons (n = 68, Fig. 1C and 1E), indicating that stimulus-synchronization is primarily used by IC neurons to encode the repetition rate of acoustic flutter. The monotonicity of the acoustic flutter-evoked response was quantified by the Spearman correlation coefficient (R value; see Methods). We defined neurons with R > 0.5 and P < 0.05 as positive monotonic (n = 97), while neurons with R < −0.5 and P < 0.05 were defined as negative monotonic (n = 18); the rest were defined as nonmonotonic (n = 49, Fig. 1D). Different from the AC having an almost equivalent proportion of positive-monotonic and negative-monotonic neurons [2], the proportion of negative monotonic neurons (11%; syn−, 4%; nonsyn−, 7%) in IC is much smaller than that of positive monotonic neurons (59%; syn+, 41%; nonsyn+, 18%, Fig. 1E).
Intriguingly, 115 out of 164 (70%) IC neurons presented either monotonic increased (positive-monotonic) or decreased (negative-monotonic) firing rates with increasing repetition rate at the flutter range (Fig.1F–1G), regardless of whether they were stimulus-synchronized or non-synchronized. Then, we fitted the temporal response of IC neurons to repetition rate at flutter range using a linear regression method and used the linear slope and mean firing rate to define their response tuning to repetition rates. Although both syn+ and nonsyn+ neurons increased firing rates with increasing repetition rates, syn+ neurons presented a larger linear fitting slope (Fig. 1H, P = 0.0048, unpaired t test) and a higher firing rate (Fig. 1I, P = 0.0029, unpaired t test) than nonsyn+ neurons. Moreover, the proportion of syn+ neurons (41%) was two times more than that of nonsyn+ neurons (18%, Fig. 1E). No significant differences in linear fitting slope (Fig. 1H, P = 0.8217, unpaired t test) and firing rate (Fig. 1I, P = 0.7437, unpaired t test) were detected between syn− and nonsyn− neurons. Interestingly, syn- exhibited better synchronization than syn+, as indicated by a larger VS (Fig. 1J, P = 0.0142, unpaired t test). Last, Syn+, syn−, nonsyn+, and nonsyn− neurons were registered into three IC subdivisions: the central nucleus (CNIC), dorsal cortex (DCIC), and external cortex (ECIC), following the same criteria as described in the previous study [12]. The majority of positive-monotonic neurons (both syn+ and nonsyn+) were predominantly localized in the CNIC (Fig. 1K). In contrast, syn-neurons primarily clustered in the DCIC, while nonsyn-neurons were mainly found in the ECIC (Fig. 1K).
The Coding of Acoustic Flutter in IC was Modulated by Sound IntensityPrevious studies have demonstrated that the majority of IC neurons monotonically increased firing rates in response to the increased sound intensity [12, 21]. So, we hypothesized that the neural coding of repetition rate at the acoustic flutter range may be affected by sound intensity. To address this question, we varied the sound level of Gaussian click trains from 20 to 70 dB SPL (Fig. 2A) and recorded IC neural responses induced by repetition rates at the flutter range. We found that the monotonicity of most IC neurons to repetition rates at the acoustic flutter range was not affected by the change of the sound intensity (Fig. 2B–2C). However, the tuning curve in response to repetition rate of some IC neurons was affected by sound intensity (Fig. 2B), and some not (Fig. 2C). Using two-way ANOVA analysis, we found that 76% of IC neurons exhibited a significantly altered tuning in response to repetition rate with varying sound levels (Fig. 2D). These results were different from those reported in the AC, that the firing rates were largely unaffected by the change of sound level [2]. To further examine how the encoding of the repetition rate at flutter range was modulated by the sound intensity, we calculated the linear fitting slope of the IC neurons in response to the repetition rate at different sound intensities (Fig. 2E, see Methods). We found that the response slopes at higher SPL were significantly greater than those at lower SPL (Fig. 2F, P < 0.0001; paired t test; Fig. 2G, ∆10 dB, P = 0.0023; ∆20 dB, P = 0.0003; ∆30 dB, P = 0.0339; ∆40 dB, P = 0.0214, paired t test). In addition, the slope difference increased with the difference between the sound intensity (Fig. 2H, P = 0.0014, F = 5.441; one-way ANOVA). Last, we examined whether syn and nonsyn neurons were affected differently by sound intensity (Fig. 2I) and found that syn neurons were affected more than nonsyn neurons by the change of sound intensity (Fig. 2I, syn, P < 0.0001; paired t test; nonsyn, P < 0.05; paired t test). In contrast to a similar percentage of SPL-sensitive and SPL-insensitive nonsyn neurons, a larger proportion of syn neurons were sensitive to the changes of sound intensity (Fig. 2J).
Fig. 2
Modulation of sound intensity on IC neural coding for acoustic flutter. SPL, sound pressure level; syn, synchronized response; nonsyn, non-synchronized response.A Schematic showing acoustic pulse trains with a repetition rate of 4–48 pps with varying sound pressure level from 20–70 dB SPL. B The examples show two IC neurons that were sensitive to changes in the sound level of acoustic flutter. Left, positive-monotonic response as a function of the repetition rate. Right, negative-monotonic response as a function of the repetition rate. Blue, 30 dB SPL; green, 50 dB SPL; orange, 70 dB SPL. The dotted line indicates the averaged spontaneous firing rate. C. The examples show two IC neurons that were insensitive to changes in the sound level of acoustic flutter. Left, positive-monotonic response as a function of the repetition rate. Right, negative-monotonic response as a function of the repetition rate. Blue, 30 dB SPL; green, 50 dB SPL; orange, 70 dB SPL. The dotted line indicates the mean spontaneous firing rate. D The proportions of sound level sensitive and insensitive IC neurons encoding repetition rate at flutter range. E The linear regression curves of an example IC neuron in response to acoustic flutter at different sound levels (blue, 30 dB SPL; green, 50 dB SPL; orange, 70 dB SPL). F The linear regression slopes of IC neurons encoding acoustic flutter at low sound levels were plotted against those at high sound levels. Purple, Δ10 dB; blue, Δ20 dB; green, Δ30 dB; orange, Δ40 dB. G Comparison of the absolute response slopes of individual IC neurons to acoustic flutter at different sound levels. paired t test, *P < 0.05, **P < 0.01, and ***P < 0.001. H The differences in linear response slope were plotted against the differences in the sound level. I The linear regression slopes of IC synchronized (syn, blue) and non-synchronized (nonsyn, purple) neurons encoding acoustic flutter at low sound levels were plotted against those at high sound levels. paired t test, *P < 0.05, ****P < 0.0001. J Proportions of IC synchronized (syn, blue) and non-synchronized (nonsyn, purple) neurons, either sensitive or insensitive to the sound level of acoustic flutter.
Then, we applied stepwise regression, a step-by-step iterative construction of a linear regression model, to examine whether the firing rate of the IC neurons was affected by the repetition rate and sound intensity of the acoustic pulse trains (see Methods). Repetition rate was introduced into the model as the first independent variable (x1), and the mean firing rate of IC neurons in response to repetition rate was used as a dependent variable (y). The regression coefficient (a) was obtained to represent the coding weight of IC neurons to repetition rate, which is named Coef-RR. To validate the regression model in repetition rate-related acoustic flutter processing, we tested this model using positive-monotonic or negative-monotonic neurons defined by Spearman correlation coefficient as shown in Fig. 1D. We found that 82 out of 86 neurons (95.3%) met the significance criteria, indicating that the computational model is highly reliable in quantifying the relative contributions of acoustic parameters in acoustic flutter processing. Subsequently, SPL was introduced into the model as the second independent variable (X2) after the repetition rate was introduced. The Coef-SPL represented the weight of SPL in acoustic flutter processing. In this model, we obtained both Coef-RR and Coef-SPL of all examined IC neurons, which represented the weights of IC neurons in encoding the repetition rate and sound intensity. An IC neuron was defined as an RR-type neuron if its firing rate was significantly affected by repetition rate only; it was defined as Both-type neuron if the firing rate was affected both by repetition rate and sound intensity; otherwise, it was defined as a None-type neuron (Fig. 3A). We found that the three types of neurons were separated and clustered in a two-dimensional space using absolute Coef-RR and Coef-SPL (Fig. 3A). None-type neurons had small absolute values of Coef-RR and Coef-SPL, which accounted for a very small proportion (Fig. 3A–3C, 5%, n = 4). Interestingly, Both-type neurons accounted for the largest proportion (Fig. 3C, n = 61) and had significantly larger absolute values of Coef-RR than Coef-SPL (Fig. 3B, P < 0.01, paired t test), suggesting that Both-type neurons had a coding bias to the repetition rate. Next, we calculated the Coef-RR and Coef-SPL of syn and nonsyn neurons separately (Fig. 3D) and found that all the None-type neurons were nonsyn neurons (Fig. 3E). The proportions of RR-type and Both-type neurons in the non-synchronized population were similar; in contrast, the number of Both-type neurons in the syn population was four times greater than that of RR-type neurons (Fig. 3E).
Fig. 3
The weight of repetition rate and sound intensity on the IC neural discharge to acoustic flutter. A The distribution and classification of Both-type, RR-type, and None-type neurons according to the absolute values of the RR coefficient (Coef-RR) and SPL coefficient (Coef-SPL), which were calculated through a stepwise regression model. Blue, Both-type neurons sensitive to both repetition rate and sound intensity; Orange, RR-type neurons only sensitive to repetition rate; Black, None-type neurons insensitive to both repetition rate and sound intensity. B The comparison of the absolute value of the RR coefficient and SPL coefficient of Both-type, RR-type, and None-type neurons. **P < 0.01, ****P < 0.0001. C The proportions of Both-type, RR-type, and None-type neurons. D The distribution and classification of IC neurons with syn (filled) and nonsyn (empty) responses, respectively, according to the absolute values of the RR coefficient and SPL coefficient, which were calculated through a stepwise regression model. E The proportions of Both-type, RR-type, and None-type neurons in IC syn and nonsyn populations. The number indicates the number of neurons. F The distribution and classification of syn+; syn−; nonsyn+ and nonsyn− neurons, respectively, according to the values of the RR coefficient and SPL coefficient. G Proportions of RR+SPL+, RR+SPL−, RR−SPL+, and RR−SPL− types in IC neurons (All) or those with syn and nonsyn responses. The number indicates the number of neurons. RR+SPL+: Positive monotonicity to both repetition rate and sound pressure level; RR+SPL−: Positive monotonicity to repetition rate and negative monotonicity to sound pressure level; RR−SPL+: Negative monotonicity to repetition rate and positive monotonicity to sound pressure level; RR−SPL−: Negative monotonicity to both parameters.
Last, to test how sound intensity is correlated with the monotonicity of IC neurons in processing of the repetition rate, we calculated the Coef-RR and Coef-SPL of syn+, syn−, nonsyn+, and nonsyn− neurons separately and used a 2-dimensional space to illustrate their correlation (Fig. 3F, only for both-type neurons). We found that the majority of neurons (72%) were located in the first quadrant, indicating the positive monotonic encoding of both repetition rate and sound intensity (Fig. 3F–3G). However, the remaining neurons (28%) were distributed in the other three quadrants (Fig. 3F). After separating syn neurons from nonsyn neurons, we found that most syn neurons clustered in the first quadrant, whereas nonsyn neurons were evenly distributed across all quadrants (Fig. 3F–3G).
The Neural Coding of Acoustic Flutter was Affected by the Click ShapeTo examine whether the IC neural coding of repetition rate in the acoustic flutter range is affected by the spectral and envelope of the Gaussian click, we varied the standard deviation (σ, sigma) of the Gaussian click to alter the frequency bandwidth and envelope of the individual click (Fig. 4A). We found that the monotonicity of most IC neurons in response to the repetition rate of acoustic flutter was also not affected by the click shape (Fig. 4B–4C). However, the tuning curve of some IC neurons to repetition rate was affected by click shape (Fig. 4B), whereas the others were not (Fig. 3C). Quantitative analyses revealed that 62% of IC neurons significantly altered their tuning curves as a function of the repetition rate in the flutter range, characterized by a higher firing rate with larger sigma values. In contrast, the remaining 38% of IC neurons showed no significant changes in their tuning properties (Fig. 4D). Next, we calculated the linear slope for individual IC neurons encoding acoustic flutter when sigma was 0.1, 0.2, and 0.4 (Fig. 4E–4F). We found no overall significant change of response slope with altered sigma (Fig. 4E–4F, 4F, P = 0.8232, F = 0.1948; one-way ANOVA). Then, we separately examined whether syn and nonsyn neurons were affected differently by sigma change (Fig. 4G). No significant difference was found (Fig. 4F, P = 0.8232, F = 0.1948; one-way ANOVA). Last, a larger proportion of syn in contrast to a smaller proportion of nonsyn neurons were sensitive to changes in sigma (Fig. 4H).
Fig. 4
The modulation of sigma on acoustic flutter-evoked response in IC neurons. Syn, synchronized response; nonsyn, non-synchronized response. A Left, the acoustic waveform (top) and spectrum (bottom) of a Gaussian click with different sigma (σ) values (0.1, 0.2, 0.4). The sigma determines the envelope and spectral component of each click. Right, acoustic flutter with varying repetition rate and sigma. Each vertical black line indicates a click. The green shaded areas indicate periods of acoustic stimulation (500 ms). B Two representative IC neurons sensitive to both repetition rate and sigma of acoustic flutter stimulation. Left, positive monotonic neuron; Right, negative monotonic neuron. The dotted line indicates the mean spontaneous firing rate. C Two representative IC neurons sensitive to repetition rate at flutter range, but not to the sigma of the Gaussian-click train. D Proportions of σ-sensitive and σ-insensitive IC neurons encoding repetition rate at flutter range. E The linear regression slopes of the IC neurons in response to repetition rate at low σ were plotted against those at high σ. Different colors represent different σ gradients. paired t test, *P < 0.05. F The difference in the linear response slope of IC neurons in response to acoustic flutter plotted against different σ gradients. G The linear regression slopes of the IC neurons in response to repetition rate at low σ were plotted against those at high σ. Different colors represent syn (blue) or nonsyn (purple) neurons. H The proportions of syn and nonsyn IC neurons are either sensitive or insensitive to the σ of the Gaussian click. The number indicates the number of neurons.
To further quantify how the repetition rate and sigma (σ) of a click affect the firing rate of the IC neurons, the stepwise regression model was used to calculate Coef-RR and Coef-Sigma for IC neurons. The IC neurons were classified into four groups: RR-type, Both-type, None-type, and Sigma-type neurons (Fig. 5A–5B). Similarly, an IC neuron was defined as RR-type if its firing rate was significantly affected by change of repetition rate only; an IC neuron was defined as Sigma-type if its firing rate was significantly affected by sigma change only; an IC neuron was defined as Both-type if the neural firing rate was affected not only by repetition rate but also by sigma change; otherwise, it was defined as a None-type neuron. We found that most IC neurons examined belong to the Both-type or RR-type neurons (Fig. 5C). Both-type neurons had significantly larger absolute values of Coef-RR than Coef-sigma (Fig. 5A–5B, 5B, P < 0.0001, paired t test). So, although the IC neural coding of repetition rate at the acoustic flutter range could be modulated by sigma, the coding bias is to represent the repetition rate over sigma. Next, we calculated the Coef-RR or Coef-Sigma of syn and nonsyn neurons separately (Fig. 5D) and found that syn and nonsyn neurons are spatially mixed. However, most of Both-type neurons were syn neurons, in contrast to similar percentage of syn and nonsyn neurons in RR-type; and all Sigma-type and None-type neurons are nonsyn neurons (Fig. 5E). Last, we further analyzed the monotonicity of Both-type neurons and plotted the Coef-RR against Coef-sigma (Fig. 5F). We found that most of Both-type neurons were distributed in the first quadrant, demonstrating a positive correlation between Coef-RR and Coef-sigma (Fig. 5F). We calculated the proportion of neurons with RR+ Sigma+; RR+ Sigma−; RR− Sigma+; RR− Sigma− in syn and nonsyn populations (Fig. 5G). We found that most syn neurons were positively modulated both by the repetition rate and Sigma (RR+ Sigma+, 72.7%, n = 16, Fig. 5G). In contrast, most nonsynaptic neurons were negatively modulated by the repetition rate, but positively modulated by the Sigma (RR− Sigma+, 66.7%, n = 2, Fig. 5G).
Fig. 5
The weight of the repetition rate and sigma of the Gaussian click on the IC discharge. Syn, synchronized response; nonsyn, non-synchronized response; syn+, positive-monotonic synchronized response; nonsyn+, positive-monotonic non-synchronized response; nonsyn−, negative-monotonic non-synchronized response. A The distribution and classification of IC neurons according to the absolute values of the (RR) coefficient (Coef-RR) and sigma coefficient (Coef-Sigma), which were calculated through a stepwise regression model. Blue, Both-type neurons sensitive to both repetition rate and Gaussian-click σ; Orange, RR-type neurons only sensitive to repetition rate; Red brown, the Sigma-type neurons only sensitive to Gaussian-click σ; Black, None-type neurons insensitive to both repetition rate and Gaussian-click σ. B The absolute value of RR coefficient and sigma coefficient in Both-type, RR-type, sigma-type, and None-type neurons. ****P < 0.0001. C The proportions of Both-type, RR-type, sigma-type, and None-type neurons. D The distribution and classification of syn (filled) and nonsyn (empty) IC neurons based on the absolute value of the RR coefficient and sigma coefficient. E The proportions of Both-type, RR-type, sigma-type, and None-type neurons in syn and nonsyn IC populations. The number indicates the number of neurons. F The distribution and classification of syn+, nonsyn+, and nonsyn- IC neurons according to the RR coefficient and sigma coefficient, which were calculated through a stepwise regression model. G The proportions of RR+Sigma+, RR+Sigma−, and RR−Sigma+ types in IC neurons (All) or those with syn and nonsyn responses. The number indicates the number of neurons. RR+Sigma+: Positive monotonicity to both repetition rate and Sigma; RR+Sigma−: Positive monotonicity to repetition rate and negative monotonicity to sigma; RR−Sigma+: Negative monotonicity to repetition rate and positive monotonicity to sigma. H–I The difference of linear response slope of IC neurons to acoustic flutter plotted against the tuning bandwidth (BW, H) or best frequency (BF, I) of IC neurons. Different colors represent different σ gradients.
To examine whether the IC neural coding of sigma in acoustic flutter was correlated with their frequency tuning properties, we plotted the difference of response slope induced by sigma change against the half tuning bandwidth (BW, Fig. 5H) and best frequency (BF, Fig. 5I) of IC neurons. We found that IC neurons with narrow tuning bandwidths were more sensitive to sigma change, exhibiting a negative correlation between the difference of response slope and tuning bandwidth (Fig. 5H, Spearman correlation coefficient, r = −0.1771, P = 0.0399); however, it was not correlated with the IC neurons’ BFs (Fig. 5I, Spearman correlation coefficient, r = −0.0799, P = 0.3659). These results indicate that the neural coding of acoustic flutter was affected by the frequency tuning properties of IC neurons. The narrower the tuning bandwidth, the stronger the influence exerted by the Gaussian click on the IC neural response.
The IC Neural Coding of Acoustic Flutter was Affected by the Irregularity of the ClickTo examine whether the IC neural coding of repetition rate in the flutter range is affected by the temporal irregularity of acoustic stimuli, we added jitter (0, 12.5%, 25%, 37.5% and 50%) into the acoustic pulse train to increase temporal irregularity (Fig. 6A). As shown by the example neuron, syn IC neurons exhibited a phase-locked response to click even when the jitter was added to acoustic pulse trains (Fig. 6B). However, the overall firing rates of some IC neurons were affected by the temporal irregularity whereas some were not (Fig. 6C). To quantify how temporal irregularity influence IC neurons’ phase-locking responses, we employed vector strength (VS) to measure the synchronization ability of IC syn neurons in response to the repetition rate with different degrees of jitter (Fig. 6D). The value of VS significantly reduced when 12.5%, 25%, 37.5% and 50% of Jitter were added to flutter stimulation (P < 0.001, one-way ANOVA). The results demonstrated a progressive decline in the neurons’ ability to synchronize with individual clicks of acoustic flutter as jitter increased. Using two-way ANOVA, we found that 50% IC neurons significantly changed their firing rate in response to the increase of jitter, whereas the firing rate of the remaining neurons did not change (Fig. 6E). Among them, 63.6% of syn neurons (n = 7) were sensitive to jitter, and a much smaller proportion of the nonsyn neurons (28.6%, n = 2) were sensitive to jitter (Fig. 6F). To further examine the coding specificity of IC neurons to repetition rate and periodicity, a stepwise regression model was applied. By using this model, we obtained both Coef-RR and Coef-periodicity of all examined IC neurons. The IC neurons in this dataset were classified into three types based on a set of significance tests in the model: RR-type, Both-type, and Periodicity-type neurons. In addition to RR-type neurons, an IC neuron was defined as a Periodicity-type neuron if its firing rate was significantly affected by temporal irregularity (jitter) only, and an IC neuron was defined as a Both-type neuron if the neural firing rate was affected not only by repetition rate but also by temporal irregularity (Fig. 6G). We plotted the absolute values of Coef-RR against those of Coef-periodicity for the three groups of I C neurons (Fig. 6H). Half of the examined IC neurons were RR-type neurons, 35% were Both-type neurons, and the rest were Periodicity-type neurons (Fig. 6I). Last, we compared the Coef-RR and Coef-periodicity for syn and nonsyn neurons (Fig. 6J). We found that all the Both-type neurons were syn neurons while all the Periodicity-type neurons were nonsyn neurons (Fig. 6J). These results were different from those of a previous AC study, showing that the firing rate of AC neurons was not affected by the temporal irregularity of acoustic pulse trains in the flutter range [2].
Fig. 6
The modulation of irregularity on the acoustic flutter evoked response in IC neurons. Syn, synchronized response; nonsyn, non-synchronized response.A Left, the schematic showing regular acoustic pulse trains spanning the range of acoustic flutter (10–45 pps) with a repetition rate varying from 4–48 pps; Right, enlarged diagrams showing irregular acoustic pulse trains with increased irregularity (Jitter: 12.5%, 25%, 37.5%, 50%). The irregular pulse train was generated by temporally shifting the time of each click in a regular pulse train, which does not affect the average repetition rate. The repetition rate of the example Gaussian click train is 16 pps. Each vertical black line indicates a single acoustic click. The green shaded areas indicate periods of acoustic stimulation, which were 500 ms. B The raster plots showing an example IC neuron in response to the repetition rate of acoustic flutter with increasing degree of irregularity (from left to right, the jitter is 0, 12.5%, 25%, 37.5%, and 50%). The green shaded area indicates the range of acoustic flutter stimulation. C The mean firing rates of IC neurons in response to acoustic flutter as a function of changing jitter during acoustic flutter. D The mean VS of IC neurons was plotted against the repetition rate with different jitters. E The proportion of jitter-sensitive and jitter-insensitive IC neurons encoding the repetition rate of acoustic flutter. F The proportions of jitter-sensitive and insensitive syn (blue) and nonsyn (purple) IC neurons. The number indicates the number of neurons. G The averaged coef-RR and coef-periodicity among different types of IC neurons. ***P < 0.001. H The distribution and classification of IC neurons according to the absolute value of the RR coefficient (coef-RR) and periodicity coefficient (coef-periodicity). I The proportions of IC neurons sensitive to both RR and jitter (both-type), only sensitive to RR (RR-type), and insensitive to both RR and jitter (none-type). J The distribution and classification of syn (filled) and nonsyn (empty) IC neurons according to the absolute values of the RR coefficient and periodicity coefficient.
Comments (0)