
Remember me



To explore the interplay between meso-endodermal patterning and tissue material-phase transitions, we quantified the joint spatiotemporal dynamics of cell fate and tissue material properties. As a readout of cell fate specification, we live-imaged the ventrolateral marginal blastoderm of zebrafish embryos labelled for membrane, nuclei, interstitial fluid and the meso-endodermal marker sebox47, starting at pluripotent stages (t = 0 min) until the onset of gastrulation (t = 120 min) (Fig. 1b,c,e). To quantify the material state of the blastoderm, we used rigidity percolation theory, a framework mapping the network representation of a material to its deformability48,49. Embryonic tissues can be abstracted as cellular networks, in which cell centroids are nodes and cell–cell contacts behave as viscoelastic bonds14,16,50 (Fig. 1c). Within this framework, the material response can be inferred by evaluating the size of the largest network cluster within which nodes have no independent movements, a collective property of the network referred to as the giant rigid cluster (GRC; Fig. 1c’). A transition in the GRC size occurs once the network crosses a critical point in the control parameter cell–cell connectivity, <k>, defined as the average number of contacts per cell48,49,51 (<kc> ≈ 4, approximately two-thirds of maximum average connectivity; Extended Data Fig. 1a,a’). Using this approach, we detect a transient spatial correlation between the specification zone and the rigid domain, where the former is secluded within the GRC at the blastoderm margin (Fig. 1c–e; t = 60–90 min). This effectively creates a physical boundary between the specifying domain and the overlaying pluripotent tissue (Fig. 1d,e, Extended Data Fig. 1e and Supplementary Movie 1). The co-occurring changes in the two macroscopic properties, meso-endodermal fate and tissue rigidity, prompted us to ask if their microscale regulators are spatiotemporally coordinated.
We explored the above question by quantifying the dynamics of the microscopic components regulating meso-endodermal fate and tissue rigidity. For meso-endodermal fate, we focused on the Nodal gradient. We performed live imaging of Smad2 and quantified the number and spatial distribution of cells with nuclear Smad2 (Fig. 1g,h, Extended Data Fig. 1f and Supplementary Movie 2). In agreement with previous reports52,53,54,55,56, we observe a short Nodal signalling range along the A–V axis, where only the first three to four most marginal cell tiers receive Nodal (closer to YSL), while cells retain the signal for ~3 h (Fig. 1h and Extended Data Fig. 1f). This pattern matches Nodal activation levels as evaluated via live imaging and quantification of the nuclear-to-cytoplasmic (N/C) ratio signal (Fig. 1i) and P-Smad2/3 immunostaining (Extended Data Fig. 1g,h), where a short-range signalling gradient is observed which steepens over time and eventually turns off.
We then quantified connectivity and GRC size along the A–V axis, to elucidate if the spatial rigidity pattern emerges as a function of spatial gradients in connectivity. We found that when the marginal tissue is still at pluripotency, the tissue is poised close to the critical point in connectivity, <kc> (Extended Data Fig. 1i). As specification progresses, connectivity increases and crosses the critical point in the most marginal cells (Extended Data Fig. 1i). The changes in connectivity seem to mainly result from a gradient in cell–cell adhesion strength (Fig. 1f,f’), which also acts as a control parameter of tissue rigidity16,57 (Extended Data Fig. 1b). Cell–cell adhesion strength can be inferred by the Young–Dupré relation, where the non-dimensional parameter α, defined as the ratio between cell–cell and cell–fluid surface tensions acting at the contact, can be derived from the angle formed at the contact edge14,16,58,59,60,61,62 (Fig. 1f and Extended Data Fig. 1b’). Quantifying α along the A–V axis revealed that, similarly to connectivity, at pluripotency, the tissue is poised close to the critical point of the relative surface tension (αc ≈ 0.866) (Fig. 1f’, box 1, and Extended Data Fig. 1j) and, during specification, the most marginal cells reduce their α values below αc, becoming therefore more adhesive (Fig. 1f’, box 3, and Extended Data Fig. 1j). Given that the critical points in α and <k> are crossed along the A–V axis, this suggests that during the rigidity transition, the initial spatial isotropy of the tissue is broken, polarizing the GRC towards the specification zone. To test if the slight gradient in the control parameters is sufficient to polarize tissue rigidity, we simulated networks and tissues with and without a gradient in <k> and α, respectively, and explored potential changes in the GRC location along the A–V axis (Extended Data Fig. 1k–n). Networks with spatially isotropic connectivity values close to criticality position the GRC at any location along the axis (Extended Data Fig. 1k,l). By contrast, under the same <k> values, a slight gradient in connectivity breaks the symmetry and polarizes the GRC towards the supercritical connectivity regions (Extended Data Fig. 1k,l and Supplementary Theory Note). The same trends are seen for α, where in silico rigidification in the presence of a slight gradient in α is sufficient to polarize the GRC along the A–V axis, concentrating its presence to the subcritical regions of α (Extended Data Fig. 1m,n and Supplementary Theory Note).
Overall, the above results show that, during patterning, the Nodal morphogen gradient arises together with a cell–cell adhesion gradient. The latter triggers a symmetry-breaking event in the initially isotropic tissue material state, correlating the spatiotemporal dynamics of tissue rigidification to specification.
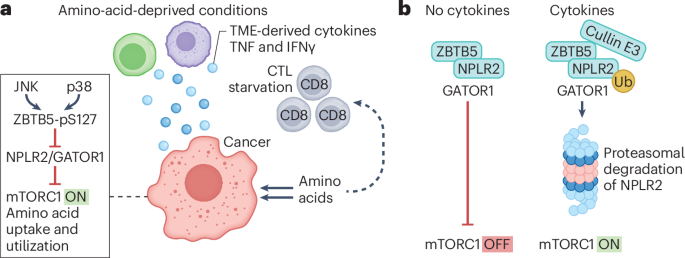
A Nodal–tissue rigidity feedback loopThe correlation between the spatiotemporal dynamics of patterning and tissue rigidity prompted us to ask whether Nodal signalling and the control parameters that drive tissue rigidification are functionally linked. First, we explored if Nodal signalling drives tissue rigidification. To this end, we quantified α, connectivity and GRC in embryos that either lack Nodal signalling (MZoep, mutant for Nodal coreceptor34,63) or have increased Nodal signalling (MZlefty1/2, mutant for Nodal inhibitors56) (Fig. 1a’). By contrast with wild-type embryos, nuclear Smad2 signal is absent in MZoep embryos, confirming that Nodal signalling is impeded. Inversely, the nuclear Smad2 region expands in MZlefty1/2 embryos, confirming that Nodal signalling is enhanced (Fig. 2a,d,e, Extended Data Fig. 2j and Supplementary Movie 2). In MZoep embryos, connectivity and α values remain close to criticality (Fig. 2b,c and Extended Data Fig. 2a), resulting in a random distribution of the GRC across the A–V axis (Fig. 2a,a’,f and Extended Data Fig. 2b). This resembles the rigidity profiles of the in silico tissues without a gradient in their control parameter (Extended Data Fig. 1k–n). Conversely, in MZlefty1/2 mutants, connectivity and α values change faster (Fig. 2b,c and Extended Data Fig. 2a), with tissue rigidity percolating throughout the entire tissue (Fig. 2a,a’,f and Extended Data Fig. 2b). Altogether, these experiments show that Nodal signalling sets the spatial rigidity pattern of the zebrafish margin by regulating cell–cell adhesion and polarizing a highly connected rigid cluster of cells towards the margin (Fig. 2f,g).
Fig. 2: Nodal triggers a tissue rigidity phase transition along the A–V axis, which feedbacks to terminate Nodal signalling. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, Exemplary 2D confocal sections at t = 90 min for wild-type, MZoep, MZlefty1/2, Opto-zGrad, MZwnt11f2/slb and MZwnt11f2/slb +Opto-RhoGEF embryos labelled with membrane–RFP (α-catenin–citrine in Opto-zGrad) and dextran-647 for interstitial fluid (a), with overlaid connectivity maps and their corresponding rigidity profiles (a’). b, Plots of GRC size as a function of connectivity over time for the conditions shown in a. Dashed lines indicate kc. The shaded arrow connects the mean connectivity and GRC values for each timepoint (n = 4 embryos wild type, n = 3 MZoep, n = 3 MZlefty1/2, n = 3 Opto-zGrad, n = 4 MZwnt11f2/slb, n = 3 MZwnt11f2/slb +Opto-RhoGEF). c, Plots of the relative surface tension α as a function of the distance from the YSL over time for the conditions shown in a, with exemplary high magnification confocal sections depicting the contact angles. Dashed lines indicate αc. Data are presented as mean ± s.e.m. (wild type: n = 12,274 contact angles, N = 8 embryos; MZoep: n = 4,003, N = 3; MZlefty1/2: n = 3,651, N = 3; Opto-zGrad: n = 2,575, N = 3; MZwnt11f2/slb: n = 6,224, N = 6; MZwnt11f2/slb +Opto-RhoGEF: n = 2,386, N = 3). d, Exemplary 2D confocal sections of embryos from the conditions described in a labelled with membrane–RFP (α-catenin–citrine in Opto-zGrad) and Smad2–GFP. The white arrowheads indicate the largest distance from the YSL with Smad2-positive nuclei. e, Plot of the distribution of nuclear Smad2-positive cells as a function of time for the conditions shown in a. Arrowheads indicate the timepoints at which the images in d were selected, corresponding to the peak of Nodal activity length scale. Data are presented as mean ± s.e.m. (n = 6 embryos wild type, n = 3 MZlefty1/2, n = 3 Opto-zGrad, n = 4 MZwnt11f2/slb, n = 3 MZwnt11f2/slb +Opto-RhoGEF). f, Quantification of GRC relative size in the specification zone (0–75 μm form YSL), for all the conditions in a, from t = 60 min to t = 120 min. Box plots show the median (centre line), the interquartile range (IQR, box: 25th to 75th percentiles) and whiskers extending to the minimum and maximum data values (n = 12 wild type, n = 9 MZoep, n = 9 MZlefty1/2, n = 9 Opto-zGrad, n = 12 MZwnt11f2/slb, n = 9 MZwnt11f2/slb +Opto-RhoGEF) g, Illustration of the Nodal–Lefty–rigidity interaction network. The time axes shown in b and c correspond to time intervals of 30 min. Statistics in f: Kruskal–Wallis followed by Dunn’s multiple comparison test, compared with wild type. All tests were two-sided. MZoep P = 0.002. MZlefty1/2 P = 0.99, Opto-zGrad P = 0.002, MZwnt11f2/slb P = 0.002, MZwnt11f2/slb +Opto-RhoGEF P = 0.99. Scale bars, 25 μm (a), 10 μm (c), 20 μm (d). n.s., non-significant.
As a next step, we asked if the collective material state affects meso-endoderm specification. To address this, we monitored Nodal dynamics using Smad2 live imaging in tissues with altered material properties. Our previous findings showed that tissue rigidity can be precisely altered by finetuning the relative surface tension α using optogenetic tools16. To this end, we used a degradation-based optogenetic system (Opto-zGrad)16,64,65 (Extended Data Fig. 2c) to maintain α close to αc and, thus, inhibit the polarization of the rigid domain. Specifically, we mildly reduced α-catenin levels by inducing its degradation in a spatiotemporally resolved manner (Extended Data Fig. 2d,e) to inhibit the decrease of α in the most marginal cells (Fig. 2a–c,f and Extended Data Fig. 2a,b). Remarkably, when examining Nodal signalling we observed that more cells and cells further away from the YSL display nuclear Smad2, indicating a shift in the length scales of Nodal signalling in the fluidized embryos (Fig. 2d, e and Extended Data Fig. 2j). Moreover, Smad2 remains active for a longer period, suggesting that the tissue material state also influences the timescale of Nodal signalling (Fig. 2e and Extended Data Fig. 2j). Importantly, the change in Smad2 dynamics is not due to indirect effects of cell–cell adhesion on other signalling pathways, such as Wnt–β-catenin66, as there is no difference in β-catenin staining between wild-type and Opto-zGrad embryos (Extended Data Fig. 2g–i). Taken together, these results show that tissue rigidity can impact Nodal signalling by restricting its spatial and temporal dynamics.
Given that the adhesion gradient and emergent rigidity observed in wild-type embryos are instructed by Nodal, we hypothesized that Nodal controls its own termination not only biochemically, via the activation of its inhibitor Lefty, but also mechanically by regulating tissue rigidity. To explore such potential negative feedback, we first asked how Nodal might influence cell–cell adhesion strength. Previous work has shown that the expression of the planar cell polarity ligand, wnt11f2, is downstream of Nodal signalling67 and that MZwnt11f2/slb mutant embryos68 display weaker cell–cell adhesion and a lower blastoderm viscosity than wild-type embryos19. We thus analysed the dynamics of cell and tissue properties in MZwnt11f2/slb mutant marginal tissues over time and space. Similarly to MZoep embryos, the MZwnt11f2/slb marginal tissue exhibits no gradient in connectivity and α along the A–V axis (Fig. 2b,c and Extended Data Fig. 2a), which remain close to their corresponding critical points, resulting in the absence of a spatially organized rigidity pattern (Fig. 2a,a’,f and Extended Data Fig. 2b). Given that MZwnt11f2/slb marginal cells are competent for meso-endoderm specification68, we investigated the effects of the absence of polarized tissue rigidity on Nodal signalling. The fluidized mutants exhibit a higher number of nuclear Smad2-positive cells which are positioned further away from the margin, and the signal is retained longer (Fig. 2d,e, Extended Data Fig. 2j and Supplementary Movie 2), thereby phenocopying the Opto-zGrad embryos. This result supports the hypothesis that Nodal is instructing changes in tissue-scale rigidity by patterning cell–cell adhesion along the A–V axis, mediated via Wnt11f2 signalling.
If the Nodal–Wnt11f2-driven rigidity negatively feeds back to terminate Nodal signalling, one would expect to rescue the Nodal signalling dynamics observed in MZwnt11f2/slb mutants, solely by re-establishing tissue rigidity. To test this, we made use of the Opto-RhoGEF tool16,69,70 (Extended Data Fig. 2f) to increase cell contractility and reduce the relative surface tension α in the most marginal cells, thereby reintroducing a cell–cell adhesion gradient in the MZwnt11f2/slb mutants. Light activation of Opto-RhoGEF in MZwnt11f2/slb embryos decreases α in a graded manner, restoring levels comparable to those in wild-type embryos (Fig. 2c). This re-introduces a gradient in connectivity (Fig. 2b and Extended Data Fig. 2a) and polarizes tissue rigidity towards the YSL (Fig. 2a,a’,f and Extended Data Fig. 2b). Strikingly, the rescue of the tissue rigidity pattern is sufficient to fully rescue nuclear Smad2 spatial and temporal dynamics, resembling those observed in wild-type embryos (Fig. 2d,e, Extended Data Fig. 2j and Supplementary Movie 2).
Altogether, these experiments demonstrate that an adhesion-driven tissue rigidity transition negatively feeds back to Nodal signalling dynamics, in a self-generated manner.
A rigidity transition restricts Nodal diffusivityThe established regulation of Nodal activity is that Nodal, as a short-range activator, induces its own expression and that of its antagonist Lefty27,33,36,37,52. Lefty, as a long-range repressor, inhibits Nodal activity by diffusing further away and binding to both Nodal and the coreceptors27,52,56,71,72. To understand how tissue rigidity regulates Nodal activity, we explored its potential involvement in the Nodal–Lefty biochemical network (Fig. 2g). Theoretical approaches indicate that the geometry of a porous environment through which particles disperse impacts diffusivity73,74,75. In the context of morphogen transport, the role of tissue architecture has been considered in poro-elastic tissues23, while experimental work proposed that the extracellular fluid structure can locally change morphogen concentration and diffusivity24,28,76. Furthermore, our recent work showed that tissue rigidification occurs in parallel to drastic changes in tissue porosity16. We therefore hypothesize that the rapid tissue reorganization occurring during tissue rigidification may directly influence Nodal kinetics within the timeframe of specification. The idea is underlined by the fact that, at αc, there is a sudden closure of small interstitial gaps between the cells via the formation of tricellular contacts16 (Extended Data Fig. 3a). This topological change leads to an abrupt collapse of the 3D interstitial fluid network (see Supplementary Theory Note Fig. SN3), in which Nodal diffuses, raising the hypothesis that a reduction in tissue porosity may restrict Nodal transport.
Quantification of tricellular contact formation during meso-endoderm specification revealed that, in wild-type pluripotent tissues with α close to αc, most of the potential tricellular contacts are open (Fig. 3a,a’). As a result, the interstitial fluid network percolates through the 3D tissue (Fig. 3a”, left). However, during specification, the tricellular contacts abruptly close when α crosses αc (Fig. 3a,a’), collapsing the interstitial fluid network (Fig. 3a’’, right). The relationship between tricellular contact formation and α is consistent for all experimental conditions (Fig. 3a–c and Extended Data Fig. 3c), supporting the claim that the topological changes leading to the rigidity phase transition occur under the same conditions at which the geometry of the cell–cell contacts triggers the collapse of the interstitial fluid network. Quantification of tissue-scale interstitial fluid fraction as a function of α revealed that porosity declines even more below αc, after the tricellular contacts are closed (Fig. 3d), suggesting that porosity may be further affected by the 3D geometrical changes of multicellular contacts. We mathematically predict that although at αc the interstitial fluid gaps vanish at tricellular points, they are still present in the central points of tetrahedral cell structures and quadrilateral gaps (Extended Data Fig. 3b and Supplementary Theory Note). By further reducing α, we find that at α ≈ 0.707 all 3D interstitial fluid gaps close in a sufficiently dense packing, reaching a minimal tissue porosity (Fig. 3d and Supplementary Theory Note). Crucially, when comparing theoretical predictions on tissue porosity against experiments, we conclude that the geometrical changes induced by the contact surface tensions can explain the changes in tissue porosity (Fig. 3d). The dynamic but drastic changes in tissue architecture occurring during specification suggest that concomitant effects on Nodal transport may arise.
Fig. 3: A rigidity–porosity transition negatively feeds back to Nodal signalling by restricting Nodal diffusivity. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a–c, Left: Exemplary 2D confocal sections of the marginal cells close to the YSL over time in wild-type (a’), MZwnt11f2/slb (b’) and MZwnt11f2/slb +Opto-RhoGEF (c’) embryos labelled with membrane–RFP, Smad2–GFP and dextran-647 for interstitial fluid. Right: Quantification of the percentage of closed tricellular contacts as a function of α for each experimental condition. Dots indicate single embryos and crosses indicate the mean for each timepoint. Timepoints are every 30 min. Black curves show simulations of the relationship between α and closed tricellular contacts. Data are presented as mean ± s.e.m. (theoretical results from ref. 16). The dotted grey line indicates αc (n = 4 embryos wild type, n = 4 MZwnt11f2/slb, n = 3 MZwnt11f2/slb +Opto-RhoGEF). a”–c”, Exemplary 3D projections of the interstitial fluid skeleton at early and late timepoints in all experimental conditions, highlighting the changes in interstitial fluid connectivity during the rigidity transition. d, Interstitial fluid fraction as a function of α, for all experimental conditions (n = 4 embryos wild type; n = 3 MZoep; n = 3 MZlefty1/2; n = 3 Opto-zGrad; n = 4 MZwnt11f2/slb; n = 3 MZwnt11f2/slb +Opto-RhoGEF; n = 3 MZoep +Opto-RhoGEF; n = 3 wild type +Opto-RhoGEF). Dashed lines indicate the critical points in α at which the tricellular gaps close and the interstitial fluid network collapses (αc1 ≈ 0.866) and all 3D multicellular gaps close and porosity is minimized (αc2 ≈ 0.707) (Supplementary Theory Note Figs. 2 and 3). e, Equations implemented to describe the biochemical feedback between Nodal and Lefty and between local Nodal levels and cell–cell adhesion. Equations 1 and 2 capture diffusion (dif), degradation (deg) and production (prod) of Nodal (N) and Lefty (L), respectively. The model was adapted from ref. 77. Equation 3 describes the accumulation of adhesion molecules as a response to Nodal levels. The plots show scaling relationships between porosity and diffusion (e’), degradation (e”) and production (e”’) rates (Supplementary Theory Note). f, Interstitial fluid fraction and Nodal range as a function of time from simulations with (grey) and without (yellow) the porosity feedback. Time is normalized to the time of maximal Nodal length scale in simulations with feedback. g–i, Quantification of Nodal range (left y axis, continuous line) and interstitial fluid fraction (right y axis, dashed line) for wild-type (g), MZwnt11f2/slb (h) and MZwnt11f2/slb +Opto-RhoGEF (i) embryos over time. Data are presented as mean ± s.e.m. (wild type: n = 6; MZwnt11f2/slb n = 4; MZwnt11f2/slb +Opto-RhoGEF: n = 3). The insets show exemplary interstitial fluid distributions at the onset and peak of Nodal signalling (n = 3 for all conditions). j–l, Left: Exemplary 2D confocal sections of the most marginal cells in wild-type (j), MZwnt11f2/slb (k) and MZwnt11f2/slb +Opto-RhoGEF (l) embryos labelled with membrane–RFP and dextran-647 for interstitial fluid. Right: MIPs of the same embryos labelled with Squint–GFP. Dashed lines indicate the YSL. j’–l’, Plots of the distribution of Squint–GFP spots as a function of the distance from the YSL for the depicted embryos in j to l. m, Plot of the Squint–GFP gradient distribution for the experimental conditions shown in j to l. Data are presented as mean ± s.e.m. (n = 5 embryos for each condition). n,o, Box plots of the fitted decay length λ and amplitude C0 for each condition. Box plots show the median (centre line), the IQR (box: 25th to 75th percentiles) and whiskers extending to the minimum and maximum data values (n = 5 embryos for each condition). p, Illustration of the Nodal–Lefty–rigidity network, showing the negative feedback fr
Comments (0)