
Remember me
Fibroblast cells have attracted significant interest because of their wide range of applications, from producing biologicals for use in transgenesis and tissue engineering. One of the key challenges in utilizing these cells is obtaining sufficient quantities while maintaining their functionality and potency. Although advanced culture techniques and strategies have significantly improved fibroblast expansion and functionality, there are still certain challenges and limitations that need to be addressed for optimal outcomes. These include accurately mimicking the mechanical, biochemical, and spatial cues that cells experience within tissues.
The extracellular matrix (ECM) is primarily built from various matrix proteins that form its main structural component, providing essential support to cells and tissues (Dzobo and Dandara 2023). ECM proteins can be categorized into two functional groups: structural ECM proteins, which provide support and integrity, and non-structural ECM proteins, also known as adhesion proteins or glycoproteins, which facilitate cellular interactions and signalling.
Foundational studies first demonstrated the importance of collagen in supporting mammalian cell cultures (Ehrmann and Gey 1956; Grobstein 1956), establishing it as a widely used structural ECM protein. Subsequent discoveries identified key adhesion proteins involved in mediating cell attachment, migration, and signalling. Fibronectin, originally termed SF/LETS protein, was shown to facilitate cell attachment and migration (Wartiovaara et al. 1974; Mautner and Hynes 1977), while vitronectin, initially known as serum spreading factor or alpha-1 protein, was found to enhance cell spreading and adhesion in serum-containing environments (Holmes 1967; Barnes et al. 1983; Hayman et al. 1983). Furthermore, laminin, discovered as a major basement membrane component, was shown to support cell differentiation and adhesion (Timpl et al. 1979). Although these ECM proteins have been well characterized in model systems, their comparative influence on primary fibroblasts from livestock species remains underexplored.
Interactions between cells and ECM elements are crucial in various biological processes because the ECM provides an array of bioactive molecules that regulate cell behaviour. Furthermore, studying the differential expression of marker genes of fibroblast identity, adhesion, oxidative stress response, and apoptosis is essential for understanding the regulatory role of ECM proteins in fibroblast function. The study of these genes will provide insights into how ECM proteins regulate cell survival and programmed cell death, further emphasizing their impact on fibroblast function and tissue homeostasis.
This study aimed to investigate the effect of structural and adhesion ECM proteins on in vitro cellular and functional responses (adhesion, proliferation, viability, colony formation, and migration) of enriched caprine adult dermal fibroblast cells (cadFibroblasts). Furthermore, we studied the influence of ECM proteins on the differential expression of fibroblast-specific, cell adhesion, antioxidant enzymes, and apoptosis-related marker genes.
This study utilized two structural ECM proteins and three adhesion (non-structural) ECM proteins, making a total of five ECM types: RTC (type 1 collagen from rat tail; Catalog#3867), collagen IV (Catalog#C5533), fibronectin (Catalog#F4759), laminin (Catalog#L2020), and vitronectin (Catalog#5051). All ECM proteins were procured from Sigma-Aldrich, Saint Louis, MO. An equal number of cells (~ 1 × 106) were cultivated onto either uncoated surfaces (control) or ECM protein-coated surfaces (treatment) under identical in vitro conditions.
cadFibroblasts from adult Barbari male goats were isolated, cultured, and enriched according to a previously described method (Pathak et al. 2025). Briefly, skin samples from the tail base of adult bucks were obtained in DPBS (DPBS; Gibco, Grand Island, NY, Catalog#14,040–117) and washed with 70% ethanol and DPBS, and the epidermis was removed using a sterile scalpel. After further rinsing with DPBS containing 10 µg/mL gentamycin (Catalog#G1264, Sigma-Aldrich, Saint Louis, MO) and 10 µL/mL antibiotic–antimycotic solution (Catalog#A5955, Sigma-Aldrich, Saint Louis, MO), the skin was trimmed into 2–3-mm2 fragments and cultured in cell culture medium (DMEM/F-12 (Catalog#D8437, Sigma-Aldrich, Saint Louis, MO) supplemented with 10 µg/mL gentamycin, 0.5% non-essential amino acids (NEAA; Catalog#M7145, Sigma-Aldrich, Saint Louis, MO), 10 µL/mL antibiotic–antimycotic solution, and 15% FBS (Catalog#10,082–147, Gibco, Fountain Drive, Paisley, UK)) at 38.5 °C with 5% CO2 for 48 h to assess fibroblast migration. Fibroblasts (cadFibroblasts) were enriched via magnetic-activated cell sorting as per the protocol described by Pathak et al. (2023). Labelled cells were separated using an MS column and eluted for subsequent experiments.
On reaching 70–80% confluence, cells were first rinsed via gentle agitation of the cell culture plate with DPBS lacking calcium and magnesium (Gibco, Fountain Drive, Paisley, UK, Catalog#14,190–136). The DPBS was then discarded, and the cell sheet (monolayer) was incubated with 1 mL of trypsin–EDTA solution (0.25%) (Catalog#25,200–072, Gibco, Fountain Drive, Paisley, UK) for 3–5 min in a CO2 incubator at 38.5 °C with 5% CO2 and humidified conditions. Cells were observed intermittently under a microscope for partial dislodgment from the surface. cadFibroblasts became rounded and detached from the plastic surface of the culture plate. Once approximately 70% of the cells were detached, 2 mL of wash medium (DMEM/F-12 10 µg/mL gentamycin, 0.5% NEAA solution, 10 µL/mL antibiotic–antimycotic solution, and 10% FBS) was added to the plate. The cells were gently collected in a 15-mL centrifuge tube and centrifuged at 2655 × g for 10 min at 25 °C to form a cell pellet. The cells were then increased in number through serial passaging in tissue culture flasks.
Immunofluorescence staining of the cadFibroblasts was conducted with adjustments to the protocol presented by Singh et al. (2022). Briefly, the fibroblast monolayer was fixed with citrate-acetone-formaldehyde, permeabilized using 0.5% Triton X-100, and blocked with 2% BSA to prevent nonspecific binding. Cells were then incubated with primary antibodies (mouse anti-vimentin (Invitrogen RV203, Catalog#MAI-06908, Waltham, MA) and rabbit anti-FSP-1 (Merck S100A4, Catalog#ABF32, Billerica MA)) and corresponding fluorescent secondary antibodies (Alexa Fluor® 488 donkey anti-mouse (Invitrogen, Catalog#R37114, Waltham, MA) and Alexa Fluor® 555 goat anti-rabbit IgG (Cell Signaling Technology, Catalog#4413S, Danvers, MA)), followed by DAPI (1 µg/mL, Sigma-Aldrich, Saint Louis, MO, Catalog#D8417) nuclear staining. Immunofluorescence imaging was conducted using a Zeiss Axiovert A1 fluorescence microscope Oberkochen, Germany.
Working solutions of each ECM protein were prepared and applied to coat culture plates following the protocol described by Singh et al. (2021). Briefly, sterile polystyrene cell culture plates were coated at specified concentrations of ECM proteins: vitronectin (100 ng/cm2), laminin (2 μg/cm2), collagen IV and RTC (10 μg/cm2), and fibronectin (5 μg/cm2) by diluting them in DPBS, adding to plates, and incubating at 37 °C for 2 h. After washing with DPBS, the plates were either used immediately or stored at 4 °C for up to 48 h, equilibrating to room temperature before use.
Cell adhesion of enriched cadFibroblasts was assessed through the trypsinization method, as described by Pathak et al. (2025). Enriched fibroblasts seeded on ECM-coated or uncoated plates were treated with 0.25% trypsin–EDTA to assess adherence. After incubation and washing, the remained adherent cells were fixed, stained with crystal violet, and examined microscopically. For quantification, crystal violet was solubilized in 1% SDS, and absorbance at 590 nm was measured using a spectrophotometer.
These assays were performed as per Pathak et al. (2023) with minor modifications. Briefly, CFU assay was conducted by plating 50,000 enriched cadFibroblast cells (P4) in ECM-coated or uncoated 6-well plates at low density. After 48 h of incubation at 38.5 °C with 5% CO2, cells were fixed and stained with crystal violet, and absorbance was measured as in the adhesion assay.
For proliferation, enriched cadFibroblasts (P4) were seeded onto ECM-coated 6-well plates (50,000 cells/well) and cultured for 10 d, with live cell counts taken every other day using an automated cell counter. Growth curves were plotted, and population-doubling time (PDT) was calculated using an online tool (http://doubling-time.com) as described by Roth (2006). Furthermore, for the scratch wound healing assay, enriched P4 cells (50,000) were seeded on ECM-coated 24-well plates and cultured to near confluence. Wounds were created using a pipette tip, and after washing, fresh media were added. Wound closure was monitored microscopically at 0, 2, 4, and 6 h, and the wound area reduction was analyzed using ZEN software.
Total RNA was extracted from enriched P4 cells using RNAiso Plus (DSS Takara, Catalog#9108, New Delhi, India). RNA concentration and purity were measured using a nanospectrophotometer (Nanodrop Lite Plus, ThermoScientific). cDNA was synthesized using a cDNA synthesis kit (DSS Takara, Catalog#6110A, New Delhi, India) and a C1000™ Thermal Cycler (Biorad, Hercules, CA), and gene expression of specific markers of fibroblast (vimentin, FSP-1), adhesion proteins (β-integrin, β-tubulin, E-cadherin), antioxidant enzymes (GPx, CuZnSOD), and apoptosis-related gene (BCL-6) was analyzed by qRT-PCR with GAPDH as the endogenous control. Reactions were performed in duplicate using TB Green® Premix Ex Taq™ (DSS Takara, Catalog#RR42LR, New Delhi India) on a StepOnePlus™ Real-Time PCR System (Applied Biosystems, Waltham, MA, Thermo Fisher Scientific, Foster City, CA), and relative gene expression was calculated using the ΔΔCt method.
The data are presented as the mean ± SEM from six independent experiments. The statistical analysis was performed using one-way ANOVA, followed by Tukey’s post hoc test. A p-value of less than 0.05 was considered indicative of a significant difference.
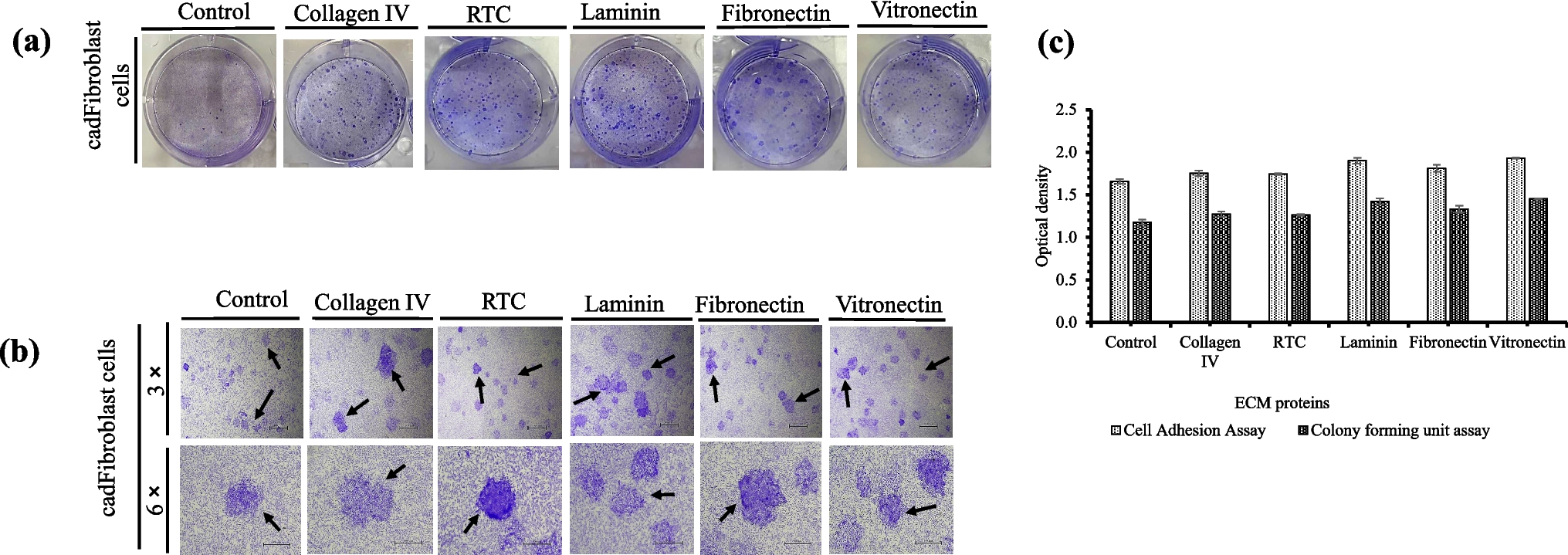
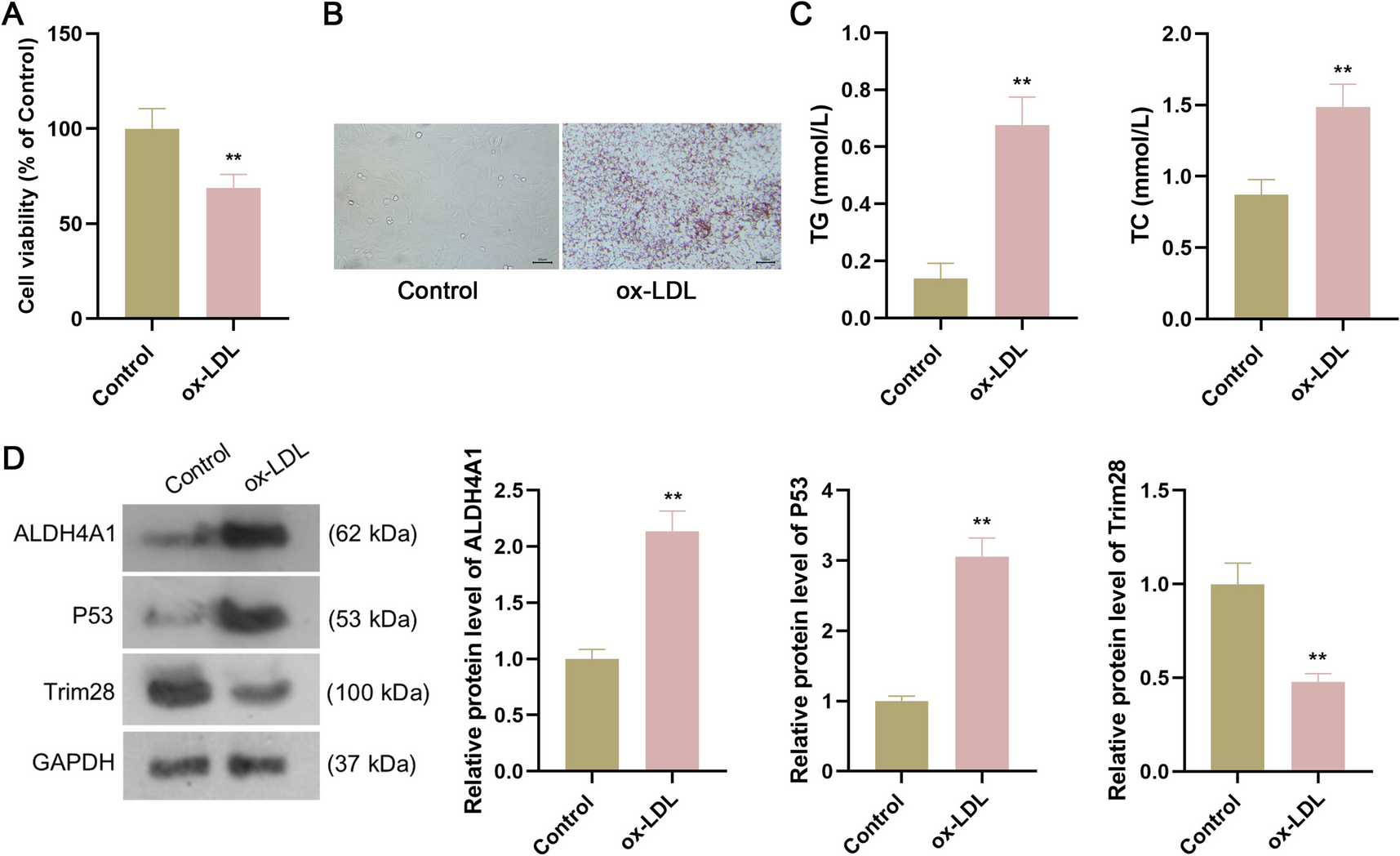
The effect of coating with structural and non-structural ECM proteins on cadFibroblast cell adhesion was investigated. Early adhesion was assessed by quantifying the number of adhered cells using crystal violet staining. Figure 1(a) shows the adhered cadFibroblast cells (stained purple) after brief (3 min) trypsin treatment, highlighting the comparison of adhesion ratios. Results indicated that laminin (1.90 ± 0.03) and vitronectin (1.93 ± 0.00) showed significantly higher cell adhesion than fibronectin (1.80 ± 0.03), while adhesion ECM proteins (laminin, vitronectin, and fibronectin) promoted greater adhesion than structural proteins like collagen IV (1.75 ± 0.03) and RTC (1.74 ± 0.00) (Fig. 1(c)). No significant differences were observed between collagen IV, fibronectin, and RTC (p > 0.05). Overall, all ECM coatings significantly enhanced cell adhesion compared to the control (1.65 ± 0.02). This trend was consistent with CFU and cell proliferation results, where faster attachment led to faster proliferation, increasing the cell number.
Figure 1.
a–c Representative pictures demonstrating the effect of different structural (collagen IV, rat tail collagen (RTC)) and adhesion (laminin, fibronectin, and vitronectin) extracellular matrix (ECM) proteins on cell adhesion and colony forming unit (CFU) efficiency. (a) The coated dishes (6-well culture plates) were seeded with 1 × 105 cells cultivated at the standard conditions described above. On attaining 70% confluence (after 48 h), the level of cell resistance to trypsinization, treatment with 0.25% trypsin–EDTA was used, and the attachment of cells to the ECM protein–coated plates versus the control cell culture plate was evaluated. Trypsin was inactivated by the addition of 2 mL of washing media, and fixation of attached cells with citrate-acetone-formaldehyde fixative solution for 30 min at RT was done. The attached cells were stained with a 1% crystal violet solution, and the optical density was compared. (b) Approximately 50,000 cells per well were cultured on 6-well cell culture plates coated with different ECM proteins in culture media with 5% CO2 at 38.5 °C in a CO2 incubator for 48 h. Large, round, and compact colonies were observed in all the groups after staining with 1% crystal violet, and the optical density was compared. Arrows indicate individual colonies. (c) Comparison of optical densities showing the effect of different ECM proteins on cell adhesion and CFU efficiency of cadFibroblasts. Optical density was measured after solubilization of stain (crystal violet) with 1% SDS solution.
In the CFU assay, isolated colonies of cadFibroblasts were observed and compared among all the treatment groups. Images of the colonies on 6-well culture plates are shown in Fig. 1(b), demonstrating differences in colony formation across the groups and the importance the impact of ECM proteins on colony formation. Absorbance measurements revealed significantly higher CFU values (p < 0.05) for laminin (1.42 ± 0.03) and vitronectin (1.45 ± 0.00) compared to fibronectin (1.32 ± 0.03). Adhesion ECM proteins (laminin, vitronectin, and fibronectin) supported greater colony formation than structural proteins like collagen IV (1.27 ± 0.03) and RTC (1.26 ± 0.00) (Fig. 1(c)). Additionally, all ECM coatings resulted in a higher number of colonies compared to the control (1.17 ± 0.02).
Cell proliferation patterns were similar across all groups up to day 2, likely due to the adaptation process as cells adjusted to the new culture conditions, which is essential for optimizing growth and viability. On day 5, an exponential growth phase was observed, with lower population-doubling times (PDT) for cells cultured on collagen IV, RTC, laminin, fibronectin, and vitronectin (50.11, 44.94, 57.49, 52.15, and 59.45 h, respectively), compared to the control (86.99 h). On day 8, inter-cellular connections were detected when cells were grown onto the laminin and vitronectin ECM proteins (Fig. 2(a)), suggesting that these adhesion proteins may be particularly effective in promoting cadFibroblast adhesion and proliferation. No stationary or declining phase occurred for cells when they were cultured with ECM proteins during the 10-d culture period, whereas a decline in cell number was noted in the control group after day 9. Overall, from day 1 to day 10, significantly higher cell count (p < 0.05) was detected in all ECM-coated groups, with lower PDTs (43.91, 42.74, 36.68, 39.96, and 38.39 h, respectively) compared to the control (55.35 h) (Fig. 2(b)).
Figure 2.
(a, b) Effect of different structural (collagen IV, rat tail collagen (RTC)) and adhesion (laminin, fibronectin, and vitronectin) extracellular matrix (ECM) proteins on proliferation of caprine adult dermal fibroblast cells (cadFibroblast). (a) Comparison of microscopic aspects of cadFibroblast with different ECM proteins from day (d) 1 to day 10 in culture. Cells on all ECM-coated plates displayed spindle-like, typical fibroblast morphology until the last day of culture, except with laminin- and vitronectin-coated plates, as cellular connections were observed after the 8th day of culture. (b) Proliferative potential of cells was determined by counting the cell number from day 1 to day 10 by an automatic cell counter. Scale bar = 100 µm.
The migration capacity of cadFibroblasts in all the groups was assessed using a scratch wound healing assay. The outcomes demonstrated a favourable effect of ECM proteins on cellular migration. For this, images were obtained and compared at different time points to evaluate wound closure. The effect of ECM proteins on cellular migration and the wound healing process was quantified by measuring the change in wound area (Table 1). A schematic diagram of the cell migration assay is shown in Fig. 3(a). As shown in Fig. 3(b), ECM proteins influenced cadFibroblast migration, affecting wound-healing capabilities. The migration rate was significantly higher (p < 0.05) for laminin and fibronectin, with fibronectin showing the highest fold change (64.77), indicating the fastest wound closure. Laminin followed with a fold change of 41.29. Additionally, vitronectin, collagen IV, and RTC showed comparable results with fold changes of 7.12, 1.92, and 3.17, respectively. Overall, all ECM proteins promoted higher migration rates than the control (1.39).
Table 1. Results presenting cell migration of fresh caprine adult dermal fibroblast cells in different extracellular matrix proteins through scratch wound healing assay. The wound area (µm2) was measured at different time intervals (immediately upon scratching (0 h), 2 h, 4 h, and 6 h) using digital photographs of selected culture areasFigure 3.
(a, b.) Migration scratch assay using caprine adult dermal fibroblast cells on different structural (collagen IV, rat tail collagen (RTC)) and adhesion (laminin, fibronectin, and vitronectin) extracellular matrix (ECM) protein–coated plates. (a) Representative pictures of scratch assay at 0, 2, 4, and 6 h with area measurement (µm2). Scale 200 µm. Approximately 50,000 cells/well were cultured on 6-well cell culture plates coated with ECM proteins in culture media with 5% CO2 at 38.5 °C in a CO2 incubator until nearly confluent to achieve growth arrest. The confluent monolayer was then scraped with a sterile small pipette. Cells migrate to cover cell cell-free area. To determine the cell migration during scratch closure, the area of each scratch was traced. and analyzed using an image processing algorithm. (b) Quantification of healing speed area (µm2) in different ECM proteins. Results represent fold change (0 to 6 h)
The relative quantification of marker gene expression for typical fibroblast, cell adhesion proteins, antioxidant enzymes, and apoptosis-related gene was conducted in all the groups. Expression analysis showed significant (p < 0.05) upregulation of fibroblast-specific and adhesion-related transcripts in cells cultured on adhesion ECM-coated surfaces, followed by structural ECM and the control group (no ECM) (Fig. 4). Likewise, the expression of GPx, CuZnSOD, β-integrin, and BCL-6 was significantly influenced (p < 0.05) by ECM proteins. Non-structural ECMs led to higher expression of antioxidant enzyme genes compared to structural ECMs. However, the expression of the apoptosis gene (BCL-6) was significantly upregulated in the control group compared to cells cultured with both structural and adhesion ECMs.
Figure 4.
Comparative evaluation of the effect of ECM proteins on the differential expression of marker genes associated with typical fibroblasts (vimentin and fibroblast-specific protein-1 (FSP-1)), cell adhesion (β-integrin, β-tubulin, and E-cadherin), antioxidant enzymes (GPx and CuZnSOD), and apoptosis (BCL-6) in enriched cadFibroblast cultured cultivated on the different ECM proteins at P-4 culture. The differential expression levels were assessed using real-time qPCR with GAPDH as an internal calibrator and DL as a control. The significant differences in other groups compared with the control group (without ECM) are presented by (a), (b), and (c) (p < 0.05). *p < 0.05 and **p < 0.01 specify ECM-dependent significant (p < 0.05) difference among the groups.
Cell culture involves extracting cells from their natural in vivo environment and growing them in an artificial in vitro setting. To ensure cells grow, function, and survive realistically, optimizing the culture environment is crucial. This includes considering factors such as the composition of the media, growth supplements, and the surface on which cells are cultured. Often, the culture conditions need to be customized to suit the specific requirements of different cell types. Consequently, the cell microenvironment is increasingly being adapted to meet the needs of particular applications by incorporating ECM into the in vitro culture environment.
In our previous study (Pathak et al. 2023), we improved the cell culture media to promote the optimal proliferation of cadFibroblasts. In the present study, we further enhanced the microenvironment of cadFibroblasts by incorporating ECM proteins into their in vitro culture conditions. Additionally, we assessed whether cell culture surface coating with ECM proteins could improve cellular morphology, adhesion, proliferation, CFU formation, migration, and differential gene expression.
In the present study, significantly higher (p < 0.05) cell adhesion and CFU ability was observed with laminin and vitronectin when compared with fibronectin, collagen IV, and RTC. This indicates that the ECM’s composition is critical in determining cell behaviour. This could be due to the reason that adhesion ECM proteins interact with a variety of integrins with high affinity, crucial for robust cell adhesion and spreading. These interactions often trigger stronger and more stable adhesion complexes compared to those formed with structural ECM proteins. This results in effective interactions with cell receptors that drive proliferation and survival, more effective cell-ECM interactions, and favourable mechanical properties provided by laminin and vitronectin (Yu et al. 2023). On the other hand, structural ECM proteins, while important for structural support and cell adhesion, do not promote proliferation and survival to the extent laminin and vitronectin do. Collagen IV, for instance, is more involved in maintaining the structural integrity of tissues rather than promoting high levels of cellular proliferation (Yu et al. 2023).
Furthermore, a statistically higher (p < 0.05) number of cells were observed with laminin, fibronectin, and vitronectin during a 10-d culture period when compared with structural ECM proteins. Moreover, the stationary and decline phase was only observed with control (no coating) during the 10-d culture period. This could be due to the absence of extracellular matrix proteins that lead to altered cell cycle dynamics and disrupted nutrient exchange. These factors could have collectively caused cells to enter a stationary phase and eventually undergo a decline phase marked by increased cell death. Proper adhesion is crucial for initiating cell proliferation. Since changes during culture could affect the cell’s attachment and proliferation, the proliferation assay was performed on cells for 0–10 d and monitored for changes in the cell number and shape.
In summary, the results demonstrated that ECM proteins differentially influenced cadFibroblast adhesion, colony formation, proliferation, viability, migration, and gene expression. Laminin and vitronectin enhanced adhesion, proliferation, and CFU formation compared to collagen and RTC. Fibronectin supported the highest wound closure, indicating a superior migratory response. Furthermore, RT-PCR showed ECM-dependent variation in gene expression. Overall, adhesion ECM proteins provided a more favourable environment for cadFibroblast growth and function than structural ECM proteins or ECM-free conditions. These findings highlight the importance of selecting suitable ECM proteins for optimizing in vitro culture systems.
Comments (0)