
Remember me

Collection and experiments on human embryos were approved by the Helsinki University Hospital Ethics Committee (diary number HUS/1069/2016). Research permission was approved by the Helsinki University Hospital Research Committee. Couples that had been treated for infertility at the Helsinki University Hospital Reproductive Medicine unit were offered a possibility to donate their cryopreserved embryos to research after termination of infertility treatments and expiration of embryo preservation timespan. Human surplus blastocysts were donated for research with an informed consent, and patients understood that donating embryos to research is voluntary. Patients were informed about the research and research methods that were used to perform the experiments. Patients were not compensated for embryo donations, and they were offered counselling. Human embryos were not created for research, nor were they manipulated for research purposes. The culture of embryos was terminated at blastocyst stage, that is, several days before the expected primitive streak formation or onset of gastrulation. We strictly follow local legislation, ethical guidelines and regulations, as well as principles laid out in the International Society for Stem Cell Research guidelines.
Day 5 and day 6 blastocysts (4AA, 4AB or 4BA) vitrified with either Kitazato Vitrification Media or Cryotech Vitrification Solutions (110, Cryotech) were warmed with Thawing Media. Cryotech strips (Cryotech) were quickly immersed in Thawing Solution at 37 °C for 1 min. The blastocysts were gently aspirated and transferred to Diluent Solution at room temperature for 3 min. Subsequently, blastocysts were transferred to Washing Solution (WS) 1 for 5 min and then to the surface of WS2 and after sinking to the bottom; this procedure was repeated, after which they were transferred to culture media (GTL, Vitrolife) and cultured in a GERI incubator for 15–16 h. All thawing solutions were from Kitazato.
Naive hES cell culture and blastoidsNaive H9 hES cells (WA09, Wicell), were converted from primed to naive pluripotency using NaïveCult Induction Kit (StemCell Technologies), and cultured on mitotically inactivated CF-1 mouse embryonic fibroblasts (Gibco) in NaïveCult Expansion Medium (StemCell Technologies) in humidified 5% O2, 5% CO2 at 37 °C. Y-27632 (10 µM; Selleckchem) was added for the first 24 h.
Blastoids were generated according to established protocols73. Naive H9 hES cells were dissociated with Accutase (Gibco), inactivated in 0.1% bovine serum albumin (BSA)–DMEM–F12, passed through a 40-µm cell strainer (Corning) and centrifuged at 300g for 5 min. Cells were plated onto 0.1% gelatin-coated dishes in NaïveCult Expansion Medium with 10 μM Y-27632 and incubated at 5% O2, 5% CO2 at 37 °C for 90 min. Non-adherent cells were collected, filtered through a cell strainer, resuspended in N2B27 basal medium and plated onto Aggrewell 400 (StemCell Technologies) plates at 6 × 104 cells per well in 500 µl N2B27 with 10 µM Y-27632, 0.3% BSA. Medium was changed 24 h after plating (day 1), and 500 µl of 2× PALLY medium was added to each well. On day 2, half of the medium was replaced with fresh 1× PALLY medium. On day 3, 500 µl of the medium was removed and 800 µl of LY medium was added. On day 4, 800 µl of the medium was replaced with fresh LY medium. PALLY medium consisted of N2B27 basal medium supplemented with 1 µM PD0325901 (MedChemExpress), 1 µM A83-01 (MedChemExpress), 10 ng ml−1 LIF (Qkine), 3 µM lysophosphatidic acid (Tocris), 10 µM Y-27632 and 1× penicillin–streptomycin. LY medium consisted of N2B27 basal medium supplemented with 500 nM LPA and 10 µM Y-27632 (Selleckchem).
hiPS cellshiPS cells were from Allen Cell Collection (Coriell). Sox2-GFP-tagged cells were used for experiments unless indicated otherwise; LaminB1-RFP and CRISPRi lines were used wherever indicated. Cells were cultured on Matrigel-coated plates in mTeSR (StemCell technologies) medium and placed in E6 basal medium (Gibco) at the onset of experiments wherever indicated, and 5 ng ml−1 TGF-β1and 100 ng ml−1 bFGF (both from StemCell Technologies) were supplemented to basal media where indicated.
Sox2-GFP hiPS cells were differentiated into mesoderm, endoderm and ectoderm lineages according to the STEMdiff Trilineage Differentiation protocol (StemCell Technologies, #05230). In brief, cells were seeded on Matrigel-coated plates 5 × 104 cells cm−2 for mesoderm induction or 2 × 105 cells cm−2 for endoderm and ectoderm induction, and were treated the following day with their respective STEMdiff media until analysed.
2D gastruloidsMicropatterns measuring 600 μm were generated using ultraviolet lithography (Alveole Primo) on plastic eight-well dishes (Ibidi) according to the manufacturer’s protocol and coated with Matrigel (1:100). Cells were dissociated with Accutase (Thermo Fischer), pelleted by centrifugation, resuspended in mTeSR1 with 10 μM Y-27632 (StemCell Technologies) and plated at 400,000 cells per well. After 1 h, the patterns were rinsed and incubated in mTeSR1 + 10 μM Y-27632 medium for 2 h, after which Y-27632 was removed. Two hours later, 50 ng ml−1 BMP4 (Miltenyi Biotech) was added.
Halotag-YAP1 and CBX2-GFP knock-in hiPS cellsFor YAP1, 0.6 pmol pUC57-Halotag-N-YAP1 plasmid (GenScript; 400 bp/ea homology arms flanking YAP1 (NM_001130145.3) start codon and Halotag coding sequence followed by GGSGGS linker) was mixed with 12.2 pmol Alt-R HiFiCas9 v3 (IDT #1081061) and 17.5 pmol single guide RNA (sgRNA; Synthego,) to transfect 226,000 hiPS cells using Nucleofector 4D 16-well cuvette, 20 μl P3 Primary Cell Nucleofector Solution (Lonza #V4XP-3032), programme CA-137. After nucleofection, the hiPS cells were cultured in rhLaminin-521 (Thermo #A29249)-coated 24-well plates in StemFlex medium (Thermo #A3349401) plus 1 μM HDR enhancer v2 (IDT #10007921) and 1× RevitaCell (Thermo #A2644501) at 32 °C overnight. Medium was subsequently changed to StemFlex plus 0.5× RevitaCell. After 3 days, cells were moved to 37 °C with StemFlex medium change every 2 days. After 16 days, cells were labelled with 1 μM Halotag Oregon Green ligand (Promega #G2802) and flow cytometry sorted (BD FASCMelody) based on fluorescence as single cells into Matrigel-coated 96-well plates. To confirm two copies of allele and no vector integration, droplet digital (dd) PCR (Bio-Rad QX200) with specific primers and probes for Halotag, AmpR (vector backbone) and reference hRPP30 was performed, further confirmed by genomic PCR of a 2.5-kb fragment, followed by Sanger sequencing.
For the mEGFP-CBX2 reporter, 400 bp/ea homology arms flanking the CBX2 (NM_005189.3) start codon were used, into which the mEGFP coding sequence was inserted, followed by a GGSGGS linker sequence (GenScript). The Kozak sequence in CBX2 was changed from GGCAGC to GGCACC to prevent CRISPR–Cas9 recutting. Then, 5 μg donor plasmid was mixed with 120 pmol Alt-R HiFiCas9 v3 (IDT #1081061) and 200 pmol sgRNA (Synthego) to transfect 800,000 hiPS cells as described above, and cultured and sorted as described above. To confirm genotype, sorted clones were first screened by 5′-junction and 3′-junction PCRs, which were further confirmed by Sanger sequencing. Clones with correct junction PCRs were then confirmed by ddPCR with one or two copies of mEGFP knock-in allele and no vector integration using mEGFP, AmpR and reference hRPP30 assay (dHsaCP1000485, Bio-Rad #10031243). Fifteen out of 32 screened clones that had one copy of mEGFP and zero copies of AmpR were selected as heterozygous mEGFP-CBX2 knock-in clones. The non-knock-in alleles in these clones were further screened by wild-type allele PCR and Sanger sequencing. For all primers and probes, see Supplementary Table 7.
Immunofluorescence and confocal microscopyHuman blastocyst stage embryos were fixed in 3.8% paraformaldehyde for 15 min at room temperature, washed three times in washing buffer (0.1% Tween20–Dulbecco’s phosphate-buffered saline (DPBS)) and permeabilized in 0.5% Triton-X-100–DPBS for 15 min at room temperature. After washing the embryos in washing buffer, embryos were incubated in Ultra Vision Protein block (Thermo Fisher Scientific) for 10 min at room temperature, followed by incubation with primary antibodies at 4 °C overnight. After three washes in washing buffer, secondary antibodies (1:500 in washing buffer) were added for 2 h at room temperature. Embryos were washed and counterstained with DAPI (1:500) and imaged in DPBS either on optical-grade plastic μ-slide eight-well chambers (Ibidi) or on glass-bottom dishes (Mattek).
Cells were fixed in 4% paraformaldehyde, permeabilized with 0.3% Triton X-100 in phosphate-buffered saline (PBS) and blocked in 5% BSA. Samples were subsequently incubated overnight in primary antibody in 1% BSA/0.3% Triton X-100/PBS, followed by PBS washed and incubation with secondary antibody in 1% BSA/0.3% Triton X-100/PBS. Cells were imaged directly after staining in PBS or mounted in Elvanol.
The following antibodies were used: OCT3/4 (Santa-Cruz Biotechnology, sc-5279; 1:1,000), Brachyury (R&D Systems, AF2085; 1:1,000), GATA6 (AF1700, RnD Systems; 1:200), NANOG (D73G4, Cell Signaling Technologies; 1:200), LAMINB1 (66095-1-Ig, Proteintech; 1:200), Pax6 (Invitrogen, #42-6600; 1:1000), SOX1 (R&D Systems, AF3369; 1:200), SOX7 (R&D Systems, AF1924; 1:1,000), YAP1 (Santa Cruz sc-101199; 1:200), p38 MAPK phosphoThr180/Tyr182 (Cell Signaling Technologies; 1:800) and CBX2 (Thermo Fisher PA-582812; 1:800). Alexa Fluor 488-, 568-, 594- and 647-conjugated secondary antibodies (all from Invitrogen) were used at 1:500 dilution.
Fluorescence images were collected by laser scanning confocal microscopy (LSM980; Zeiss) with Zeiss ZEN Software (Zeiss ZEN v.3.7), or with Andor Dragonfly 505 spinning disk confocal (Oxford Instruments) equipped with 488-nm and 546-nm lasers, and Zyla 4.2 sCMOS camera using 40×, 63× or 100× immersion objectives and Fusion software (v.2.3.0.44, Andor). Images were acquired at room temperature using sequential scanning of frames of 1-µm confocal planes (pinhole 1).
Live imagingSamples were mounted on a Andor Dragonfly spinning disc confocal (Oxford Instruments) equipped with an environment chamber (5% CO2 and 37 °C). Images were acquired with Fusion software (v.2.3.0.44, Andor). For nuclear volume measurements LaminB1-RFP hiPS cells were live imaged for 10 min (growth factor experiment) or 40 min (cell compression experiments) acquiring full z-stacks from the same colony for each timepoint using a 63× water immersion objective. For 2D gastruloid micropatterns, cells were imaged with a 20× air objective, the z-step was 0.75 µm and images were captured every 1 h. For nuclear envelope fluctuations, cells were imaged with a 63× oil immersion objective using a high frame rate acquisition mode (150 ms per frame) for 5 min. For CBX2 dynamics, mEGFP-CBX2 hiPS cells were imaged with a 63× water immersion objective, focusing on single focal plane at the widest point of the nuclei. Images were obtained at at 5 min per frame.
Segmentation and image analysisNuclear volume quantificationVolumes were quantified using ImageJ74. Four-dimensional live imaging videos were filtered using a 3D median filter and subsequently bleach corrected using simple ratio correction. Individual nuclei were identified manually and marked with an oval selection stored to a region of interest (ROI) manager. Seeds were then expanded using the Limeseg-plugin75. The segmentation was manually supervised to ensure complete segmentation.
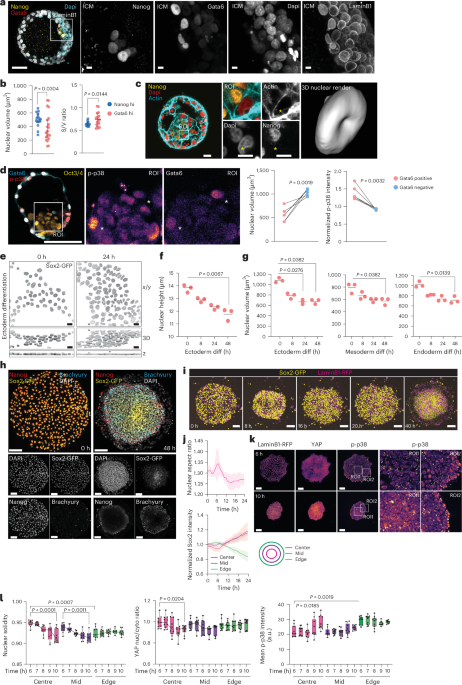
For human embryos, the segmentation was performed similarly with the following modifications. Fixed immune-stained embryos were imaged using a 40× oil immersion objective, and cells were identified as ICM on the basis of Nanog expression. ICM cells with high expression of Nanog or Gata6 were then manually identified and segmented on the basis of LaminB1 signal. For volume calculation in three-lineage differentiation experiments, Sox2-GFP-expressing hiPS cells were plated on glass-bottom 35-mm gridded bottom dish (Ibidi) to image the same colonies of cells across multiple timepoints. A complete z-stack was imaged using a Nikon eclipse Ti2 inverted microscope mounted with a CSU-W1 spinning disk microscope (60× oil immersion lens, numerical aperture (NA) 1.49) immediately before starting the differentiation protocol. The same colonies were imaged 8 h, 24 h and 48 h after the addition of differentiation media. Then, 3D segmentation of nuclei (outlined by Sox2-GFP expression) was performed using a custom Cellpose (v.2.2.2) model76,77 to generate 3D masks. The 3D masks were converted to 3D ROIs using the 3D Suite plugin78, after which nuclear volumes was quantified from 3D ROIs using the same plugin.
Nuclear envelope fluctuationsImages were corrected for bleaching (Bleach Correction function of ImageJ) and linear rotational drift (Stackreg; ImageJ). After corrections, the nuclear edge position was recorded as a function of time at different positions along the NE by drawing a line perpendicular to NE at multiple locations per nucleus. Fluctuations were calculated by measuring the standard deviation of the position of the NE from its mean position46. Actinomycin D (10 µM)/2-desoxyglucose (5 mM), Cytochalasin D (200 nM), Calyculin A (5 nM) and Nocadozole (400 nM; all from Sigma) were added where indicated. SPY650-FastAct (SpiroChrome) and MemGlow 488 (Cytoskeleteton) were used to visualize actin and the plasma membrane.
CBX2 cluster analysisImages were obtained using Zeiss LSM980 plus or Airy Scan imaging mode and subsequent deconvolution. Individual cell nuclei were segmented from the images using custom Cellpose (v.2.2.2) model76,77. CBX2 clusters were detected with Cellpose using a custom-trained model. The number of aggregates per cells were counted using ImageJ. For localization at the nuclear periphery nuclei Cellpose masks were dilated 400 nm isotropically and eroded 20 times for 150 nm to generate thin consecutive masks using ImageJ. The thin masks were used to measure spatially resolved mean intensities of CBX2 and DAPI from the nuclear periphery from background-subtracted images. Each measurement was then divided by the mean total nuclear intensity of CBX2.
For condensate detection, live imaging videos were preprocessed with the Noise2Void noise reduction algorithm79 and segmented using a Cellpose custom-trained model. The nuclear outlines were tracked on the basis of the overall CBX2 nuclear signal using TrackMate Cellpose adaptation80. For relative aggregation, CBX2 unprocessed videos were processed with ImageJ bleach correction plugin using the histogram matching algorithm. Relative aggregation was calculated for each tracked cell from background-subtracted videos by measuring the summed grey values within the condensate area and dividing them by the summed grey values of the entire nuclear area.
Quantification of nuclear intensities of transcription factorsMean intensities were measured from images using a nuclear mask generated from LaminB1-RFP, DAPI or Sox2-GFP using ImageJ74. The N/C ratio for YAP staining was calculated for each cell by dividing the nuclear mean intensity by the cytoplasmic mean intensity.
Mechano-osmotic perturbationsCompression was performed with a modified version of a previously published cell confiner system81. In brief, suction-cup-bound coverslips were applied to 2D iPS cell colonies using a controlled pressure pump (Elveflow). Compression height was controlled by 5-µm polystyrene bead spacers.
For the western blotting, phosphoproteomics and sequencing experiments cells were axially compressed using polystyrene block custom manufactured to fit 6-cm or 10-cm cell culture dishes. The blocks were preheated to 37 °C and placed on cells, ensuring a homogeneous layer of culture medium between the block and the cell layer.
Hyperosmotic shock was induced using 0.5 M sucrose in media. For CBX2 live imaging in hypertonic conditions, hypertonic buffer was diluted 1:20. Hypo-osmotic shock was induced using preheated mQH2O that was added to the medium to achieve a final 1:4 mixture.
AFMAFM force spectroscopy measurements were performed using JPK NanoWizard 2 (Bruker Nano) mounted on a Nikon Eclipse Ti inverted microscope and operated with JPK SPM Control Software v5. Measurements were performed at 37 °C. Triangular non-conductive silicon nitride cantilevers (MLCT, Bruker) with a nominal spring constant of 0.01 N m−1 were used for force spectroscopy measurements with the probe tip positioned directly over the nucleus. For all indentations, forces of up to 3 nN were applied and the velocity of indentation was kept constant at 2 µm s−1. Force maps of 4 µm2 with resolution 2 × 2 pixels were used to perform technical replicates of nuclei indentation, and all valid curves were analysed. Before fitting the Hertz model to obtain Young’s Modulus (Poisson’s ratio of 0.5), the offset was removed from the baseline signal, the contact point was identified and cantilever bending was subtracted from each force curve. All analysis was performed with JPK Data Processing Software (Bruker).
nucGEMShiPS cells were transfected using X-tremeGENE (Sigma) according to the manufacturer’s instructions. Colonies were imaged 36 h after transduction on a Nikon Eclipse Ti Eclipse microscope mounted with a Yokogawa CSU-W1 spinning disk unit using 405-nm and 488-nm lasers, a 63X CFI Apo 60×/NA 1.49/0.12 total internal reflection fluorescence microscopy objective and an ET525/36 m emission filter (Chroma). Images were acquired using 80% power from single focal plane at 100-ms intervals, 1 × 1 binning and 16-bit pixel depth. The High-Throughput Image Processing Software platform82 was used to track particles, and custom Python pipelines were used to calculate aggregate mean square displacement, effective diffusivity and the diffusive exponent83. Single-particle tracks were generated with the Napari GEMspa plugin84.
Multiome sequencing and analysisCells were compressed as described above, after which single nuclei were processed using the Chromium Next GEM Single Cell Multiome ATAC + Gene Expression kit. Library preparation were performed according to the manufacturer’s protocol. Two biological replicates were prepared for each condition and sequenced using the Chromium Single Cell Multiome platform (10x Genomics). All replicates were quality controlled and analysed separately to ensure reproducibility, after which all conditions were pooled and analysed together, treating each cell in the total pool as a biological replicate. Initial transcript count and peak accessibility matrices were obtained with Cell Ranger Arc (v.1.1.2).
Raw counts were subsequently imported into Python (3.8) as AnnData (0.8.0) objects. Cells with more than 25% mitochondrial RNA content were removed. Doublet prediction on single-cell (sc)RNA-seq data was performed using scrublet (0.2.3). Raw scRNA-seq counts were normalized using scran (1.22.1) with Leiden clustering input at resolution 0.5. scRNA-seq and scATAC-seq data were integrated as MuData objects (0.2.2) using Muon (0.1.2). Raw scATAC-seq counts were filtered for noise, and subsequently Term Frequency–Inverse Document Frequency was transformed using muon.atac.pp.tfidf with a scale factor of 10,000.
scRNA-seq data were further processed using scanpy (1.8.2): for 2D embedding, the expression matrix was subset to the 2,000 most highly variable genes (sc.pp.highly_variable_genes, flavour ‘seurat’). The top 50 principal components were calculated and served as the basis for k-nearest-neighbour calculations (sc.pp.neighbors, n_neighbors = 30), which were used as input for uniform manifold approximation and projection (UMAP; https://doi.org/10.48550/arXiv.1802.03426) layout (sc.tl.umap, min_dist = 0.3).
scATAC-seq data dimensionality was reduced to 50 components using muon.atac.tl.lsi, and embedded in a 2D UMAP (sc.tl.umap) on the basis of k-nearest-neighbour calculation (sc.pp.neighbors, n_neighbors = 20).
Gene signatures for cell cycle85, apoptosis (BCL2L1, CASP9, CYCS, IL1A, PIK3CG, TNFRSF10D, FADD, BIRC3 and FAS), starvation (ATG5, ATG12, ULK2, GABARAPL1, PRKAA2, BNIP3, TRIB3, DDIT3, HSPA5 and SERPINB3), hypoxia (HIF1A, HIF3A, EPAS1, ARNT, PCNA, ADM, CCND1, CA9 and GLUT1) and necrosis (BIRC3, FAS, DNM1L, GSDME, IPMK, MLKL, RBCK1, TICAM1 and YBX3) were scored using the scanpy ‘sc.tl.score_genes’ function. Pychromvar (0.0.4) was used to interrogate transcription-factor accessibility. Pseudobulk differential expression analysis was performed with pyDESeq2 (0.4.8). Active enhancers and gene regulatory networks were inferred using SCENIC+50 (pyscenic 0.11.2) following documentation. In brief, topic modelling was performed with 16 topics after evaluating different Latent Dirichlet allocation models, and binarized using the Otsu method. Condition annotations were added to the metadata alongside topics. Accessibility matrices were imputed and normalized, and highly variable features and condition-specific accessibility were calculated. Custom motif and ranking databases were created using ‘create_cisTarget_databases’ based on the observed scATAC-seq peaks. Motifs were obtained from the SCENIC+ ‘v10nr_clust_public’ collection. After running SCENIC+, eRegulons underwent standard filtering and scoring.
H3K27ac Cut&Tag and analysisCut&Tag was performed using Complete CUT&Tag-IT Assay Kit (Active Motif). Three biological replicates were collected from 10-cm dishes by brief incubation at 37 °C in 0.05 M EDTA in Hank’s Balanced Salt Solution and attached to Concanavalin-A-conjugated magnetic beads (ConA beads, Polysciences) before overnight binding of primary antibody H3K27ac (Abcam, ab4729). Samples were then incubated with secondary antibody and CUT&Tag-ITAssembled pA-Tn5 Transposomes, after which tagmented DNA was purified, PCR amplified using a i5/i7 indexing primers, cleaned using SPRI beads and pooled into an equimolar library for sequencing on an Illumina HiSeq4000 sequencing platform.
Unmapped paired-end reads were trimmed to remove adapters and poor-quality sequences using fastp v.0.23.2 (--detect_adapter_for_pe). Paired-end reads were mapped to GRCh38 and Drosophila melanogaster (GCF_000001215.4_release_6_plus_iso1_mt) reference genomes using bwa-mem2 v.2.2.1 with default settings. To define active enhancers and promoters, we performed peak calling using the callpeak function (‘-f BAMPE -g hs -q 1e-5 --keep-dup all --nomodel’) from MACS2 (2.2.7.1)86. PCR duplicates were removed using sambamba (v.1.0.1)87, and the IgG-negative control from the CBX2 Cut&Run experiment was used for estimating background. The final set of H3K27ac-enriched genomic regions included a union of peaks from all experimental conditions that were detected in at least two biological replicates, with peaks closer than 1 kb merged together and peaks overlapping with the ENCODE blacklisted genomic regions discarded88. All H3K27ac peaks were further classified into active promoters (n = 10,728) and putative active enhancers (n = 15,531) based on the overlap with transcription start site (TSS) ± 1 kb regions of hg38 UCSC refGene and protein-coding GENCODE v38 genes. To link putative active enhancers with their potential target genes, we used the closest function from BEDTools (v.2.31.1)89 to find the nearest UCSC refGene gene (TSS ± 1 kb region).
Following PCR duplicate removal, biological replicates (two replicates for pluripotency and hypertonic stress conditions, three for all others) from the same condition were spike-in normalized and merged together90 and then used to generate coverage tracks using bamCoverage from deepTools (v.3.5.4)91 (‘-bs 1 –ignoreDuplicates’). BigwigCompare from deepTools (‘--skipZeroOverZero --operation log2 --fixedStep -bs 50’) was used to generate the differential H3K27ac enrichment tracks. To perform metaplot and heatmap analysis of the mean read density at regions of interest, we used computeMatrix and plotProfile/plotHeatmap from the deepTools suite.
Significant changes (p-adj <0.05, |FC| >1.5) in H3K27ac enrichment at active enhancers following different treatments were identified using the DESeq2 R package (v.1.42.1)92,93. In brief, multiBamSummary from deepTools (‐‐outRawCounts) was used to obtain counts of hg38 mapped reads at target regions of interest across different conditions, following PCR duplicate removal. Read counts from the spike-in dm6 genome at dm6 refGene genes were used to calculate size factors for spike-in calibrated DESeq2 analysis90. To visualize an overlap of active enhancers decommissioned following compression (a union of peaks showing a significant loss of H3K27ac in the compressed pluripotency and basal conditions, n = 3,694) and hyperosmotic shock (n = 5,496), we used UpSetR (v.1.4.9) and VennDiagram (v.1.7.3) R packages. Gene Ontology (GO) term analyses were carried out using Enrichr94.
TTseq and analysisCells were labelled with 500 µM 4‐thiouridine for the last 10 min of the indicated 30-min treatments, after which cells were collected and lysed in TRIzol. ERCC spike-ins (00043, 00170 and 00136) were added to lysates, after which RNA was isolated, fragmented and biotinylated with EZ-link HPDP-biotin (Thermo Fischer). Biotinylated nascent RNAs were purified with streptavidin-conjugated magnetic beads (µMACS; Miltenyi), and libraries of total and biotinylated RNA were prepared with Illumina TruSeq total RNA kit. Libraries were quantified using the KAPA Library Quantification Kit and sequenced with Illumina NextSeq 500 using the High Output Kit v.2.5 (150 cycles, Illumina) for 2 × 75-bp paired-end reads. Trimmed reads were aligned to the human genome assembly (hg38) using STAR2.4.295. For coverage profiles and visualization, reads were uniquely mapped, destranded, antisense corrected and normalized with size factors calculated from DESeq292.
To quantify changes, spike-in calibrated differential analysis was performed92. DESeq2 with size factors estimated using ERCC spike-ins (00043, 00170 and 00136) was used for differential expression. Raw read counts following PCR removal were obtained for a set of hg38 MANE RefSeq genes (v.1.3)10 using the featureCounts function (‘isPairedEnd = TRUE, countReadPairs = TRUE, useMetaFeatures = TRUE, strandSpecific = 1’) from the Rsubread package (v.2.16.1). log2FCs following shrinkage using the original DESeq2 estimator7 were visualized using custom R scripts and ggplot2.
Bulk RNA-seq and analysisTotal RNA was isolated using the NucleoSpin RNA Plus kit (Macherey-Nagel). After quantification and quality control using Agilent 2200 TapeStation, total RNA amounts were adjusted and libraries were prepared using the TruSeq Stranded Total RNA kit with Ribo-zero gold rRNA depletion (Illumina). RNA-seq was carried out on Illumina NextSeq 500 using the High Output Kit v.2.5 (150 cycles, Illumina) for 2 × 75-bp paired-end reads.
Raw FASTQ files were adapter-trimmed and quality-filtered using fastp (0.23.2) using default settings and the ‘—detect_adapter_for_pe’ flag. Filtered reads were mapped to the GRCh38 reference genome merged with ERCC92 spike-in sequence reference using STAR (2.7.10a) (--outSAMstrandField intronMotif --outFilterIntronMotifs RemoveNoncanonical --outFilterMultimapNmax 1 --winAnchorMultimapNmax 50 --outFilterType BySJout --alignSJoverhangMin 8 --alignSJDBoverhangMin 1 --outFilterMismatchNmax 999 --outFilterMismatchNoverReadLmax 0.04 --alignIntronMin 20 --alignIntronMax 1500000 --alignMatesGapMax 1500000 --chimSegmentMin 15 --chimOutType WithinBAM --outSAMattributes All). Gene expression counts were generated from mapped reads using Gencode v35 and ERCC92 annotations with htseq-count (2.0.3).
PCA and differential gene expression analysis were performed in R (4.1.2) using DESeq2 (1.34.0) assuming negative binomial distribution of read counts (Wald test). GO term and transcription factor binding analyses were carried out using Enrichr/ChEA394.
Phosphoproteomics and analysishiPS cells in pluripotency media were subjected to compression or hypertonic shock for 5 min, after which cell lysates were collected in 6 M guanidinium chloride buffer supplemented with 5 mM Tris(2-carboxyethyl)phosphine, 10 mM chloroacetamide in 100 mM Tris-HCl. Samples were then boiled at 95 °C for 10 min, sonicated at high performance for 10 cycles (30 s on/off) using Bioruptor Plus Ultrasonicator (Diagenode) and spun down for 20 min at room temperature at 20,000g. Supernatants were then trypsin-gold digested (Promega Corp., V5280) overnight at 37 °C. Digested samples were acidified and peptides were cleaned with custom-packed C18-SD Stage Tips. Eluted peptides were vacuum dried at 30 °C, and phosphopeptides were enriched using the 3 mg/200 ml Titansphere Phos-TiO kit (GL Sciences) according to the manufacturer’s instructions. After elution, phosphopeptides were vacuum dehydrated for 2 h at 30 °C and cleaned with custom-packed C18-SD. Ultimate 3000 ultra-high-performance liquid chromatography in conjunction with high-pH reversed-phase chromatography was used to separate and fractionate a total of 1 mg of tryptic peptides, from which eight fractions were collected.
All samples were analysed on a Q Exactive Plus Orbitrap mass spectrometer that was coupled to an EASY nLC (both Thermo Scientific). Peptides were loaded with solvent A (0.1% formic acid in water) onto an in-house packed analytical column (50 cm, 75 µm inner diameter, filled with 2.7-µm Poroshell EC120 C18, Agilent). Peptides were chromatographically separated at a constant flow rate of 250 nl min−1 using the following gradient: 3–5% solvent B (0.1% formic acid in 80 % acetonitrile) within 1.0 min, 5–30% solvent B within 121.0 min, 30–40% solvent B within 19.0 min, 40–95% solvent B within 1.0 min, followed by washing and column equilibration. The mass spectrometer was operated in data-dependent acquisition mode. The MS1 survey scan was acquired from 300–1,750 m/z at a resolution of 70,000. The top ten most abundant peptides were isolated within a 1.8-Th window and subjected to Higher-energy collisional dissociation (HCD) fragmentation at a normalized collision energy of 27%. The AGC target was set to 5 × 105 charges, allowing a maximum injection time of 55 ms. Product ions were detected in the Orbitrap at a resolution of 17,500. Precursors were dynamically excluded for 25.0 s.
Raw data were processed with MaxQuant (v.2.4.0)96 using default parameters against the Uniprot canonical Human database (UP5640, downloaded 4 January 2023) with the match-between-runs option enabled between replicates and phosphorylation at S, T and Y defined as variable modifications. Results were loaded into Perseus (v.1.6.15)96, and contaminations and insecure identifications were removed. Remaining identified phosphosites were filtered for data completeness in at least one replicate group, and remaining data were normalized by median subtraction for each individual samples. After sigma-downshift imputation using standard settings, significantly changed sites were identified using both one-way analysis of variance (ANOVA) and false discovery rate-controlled t-tests and annotated using PhosphoSitePlus97. One-dimensional enrichments were performed on site abundance differences. ANOVA-significant site abundances were z-scored and hierarchically clustered using standard settings. GO term analyses were carried out using Enrichr94.
Western blottingCells were rinsed in PBS, suspended in lysis buffer (50 mM Tris-HCl buffer (pH 8.0), containing 150 mM NaCl, 1% Triton X-100, 0.05% sodium deoxycholate, 10 mM EDTA, protease and phosphatase inhibitors) and cleared by centrifugation. The lysates were then reduced in Laemmli sample buffer at 95 °C, separated by polyacrylamide gel electrophoresis in the presence of SDS and transferred onto polyvinylidene fluoride membranes. Membranes were blocked with 5% milk powder in Tris-buffered saline containing 0.05% Tween (TBS-Tween) for 1 h at room temperature, after which primary antibodies were added in 5% BSA, TBS-Tween and incubated overnight at 4 °C. The membranes were subsequently washed in TBS-Tween, after which secondary horseradish peroxidase-conjugated antibodies (Bio-Rad) were added in 5% milk powder in TBS-Tween and incubated for 30 min at room temperature. Antibody binding was detected by chemiluminescence (Immobilon Western, Millipore) using the Bio-Rad ChemiDoc Imaging System. Antibodies used: LaminB1 (Cell Signaling 9087; 1:1,000), p38 MAPK phosphoThr180/Tyr182 (Cell Signaling 4511, 1:1,000), p38 MAPK (Cell Signaling 9212, 1:1,000), p44/42 MAPK phosphoErk1/2 (Cell Signaling 4376, 1:2,000), p44/42 MAPK Erk1/2 (Cell Signaling 4695, 1:1,000) and RNApol2 PS2 (Abcam ab5095; 1:5,000).
RT–qPCRCells were treated as indicated with media conditions, compression, YAP inhibitor verteporfin (200 nM) and ERK inhibitor (1 µM). RNA was isolated using the NucleoSpin RNA plus Kit (Macherey-Nagel). RNA quality was assessed using the high-sensitivity RNA Screen Tape Analysis (TapeStation, Agilent), and subsequently, 500 ng of RNA was reverse transcribed applying the SuperScript IV VILO Master Mix (Thermo Fisher Scientific) following the manufacturer’s instructions. External RNA Controls Consortium RNA Spike-in Mix (Thermo Fisher Scientific) was added to samples as reference to ensure validity of normalization. PCR was performed with PowerUp SYBR Green Mastermix (Thermo Fisher Scientific) using QuantStudio5 Real-Time PCR System (Thermo Fisher Scientific). Gene expression was quantified using the ΔΔCt method with normalization to LaminB1. For primers, see Supplementary Table 7.
CBX2 Cut&Run and analysesCells were compressed from 30 min in basal or pluripotency medium followed by 24 h recovery in the same media condition. Uncompressed cells in the respective media were used as controls (two biological replicates per condition). CBX2 cut&run was performed using the CUTANA ChIC Cut&Run Kit (EpiCypher) using the manufacturers’ instructions. In brief, 50,000,000 cells per replicate were collected and absorbed with activated ConA beads, followed by incubation with 0.5 µg of CBX2 antibody (mAb #25069, Cell Signaling) overnight at 4 °C. Chromatin was then digested and released, followed by DNA purification and library preparation using the CUTANA Cut&Run Library Prep Kit (EpiCypher). After library quantification and quality control using Agilent 2200 TapeStation, DNA sequencing was carried out on Illumina NextSeq 500 using the High Output Kit v2.5 (150 cycles, Illumina) for 2× 75-bp paired-end reads.
Raw FASTQ files were adapter-trimmed and quality-filtered using fastp (0.23.2) using default settings and the ‘—detect_adapter_for_pe’ flag. Filtered reads were mapped to the GRCh38 and Escherichia coli (ASM886v2) reference genome using bwa-mem2 (2.2.1). Mapped reads were quality-filtered using a cut-off of 3. Scaling-factor normalized tracks were generated according to Zheng et al. (https://doi.org/10.17504/protocols.io.bjk2kkye). Regions were blacklisted using published CUT&RUN blacklists98.
Coverage profiles for IgG and CBX2 were generated using MACS2 (2.2.9.1). Normalization to IgG was performed using MACS2 ‘bdgcmp’ and the ‘qpois’ method, and peak calls were obtained using MACS2 ‘bdgbroadcall’ (-c 5 -C 2 -g 1000 -G 2000 --no-trackline). Peaks were filtered for noise using reproducibility (called in >1 sample of the same condition), minimum peak size (>1,000 bp), and minimum mean CBX2 signal in one condition (>0.075).
Aggregate statistics for each sample were calculated on replicate-merged tracks for peaks overlapping known genes (Gencode v44). Mean peak signal was scaled to the pluripotency medium as reference condition, and log2-transformed for symmetry. Hierarchical clustering was performed using the Ward method and Euclidian distance metric (scipy 1.11.4); clusters were chosen with a tree distance cut-off of 7.
CRISPRi-mediated silencing of CBX2One microgram of Alt-R CRISPR sgRNA (IDT; 5′-CCCGGCAGCCAGCCCGACCG) targeting the CBX2 promoter was mixed with 1 × 105 WTC-dCas9-TagBFP-KRAB hiPS cells (Coriell #AICS-0090-391) and electroporated using the Neon Transfection System (Thermo Fisher Scientific), using a regime of 1,400 V, 10 ms and two pulses. After electroporation, hiPS cells were cultured in Matrigel coated plate with mTeSR medium plus 25 μM Y-27632 ROCK inhibitor (Tocris #1254). After 24 h, the medium was changed to mTeSR without Y-27632 with fresh medium change daily. Ninety-six hours after sgRNA delivery, a subset of cells was fixed and immunostained for CBX2 to assess silencing efficiency. The remaining cells were transferred to E6 basal medium and cultured for an additional 96 h with daily medium changes.
Statistics and reproducibilityStatistical analyses were performed using GraphPad Prism software (GraphPad, v.9) or in R (v.4.2.2). Statistical significance was determined by the specific tests indicated in the corresponding figure legends. Only two-tailed tests were used. In all cases where a test for normally distributed data was used, normal distribution was confirmed with the Kolmogorov–Smirnov test (α = 0.05). All experiments presented in the Article were repeated in at least three independent replicates. No statistical method was used to predetermine sample size. No data were excluded from the analyses. The experiments were not randomized as groups were assigned by treatments. As data analyses where automated, investigators were not blinded for outcome assessment.
Reporting summaryFurther information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Comments (0)