
Remember me



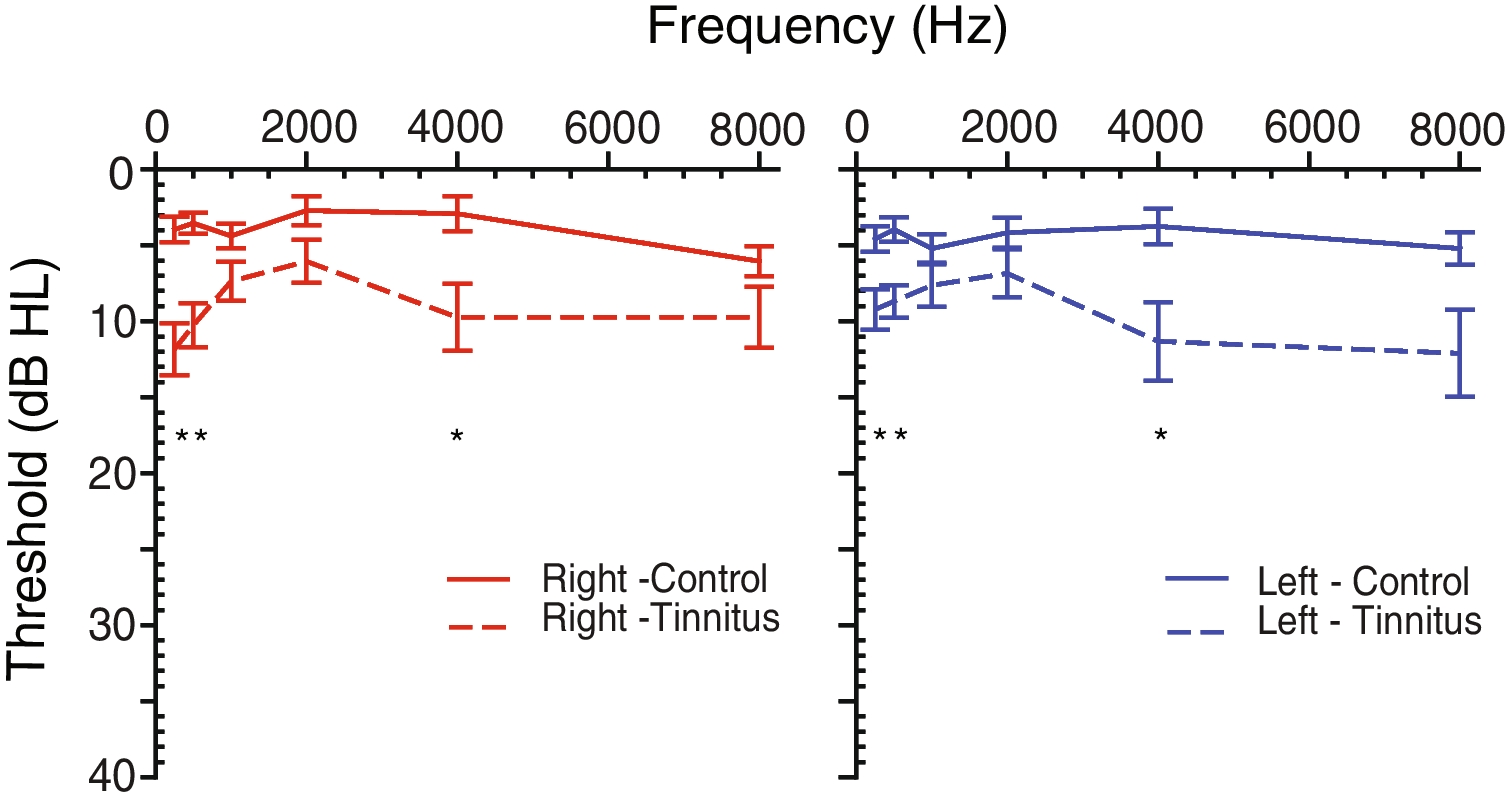

Approximately half of the people aged 60, and increasing in prevalence from age 60 onwards, will suffer from age-related auditory dysfunction, both when self-reported and measured audiologically [1]. Classically, patients present with two main symptoms of auditory dysfunction: either difficulty in detecting sounds or in perceiving sounds [2]. While sound detection often manifests as not being able to hear sounds of a low amplitude as a result of disrupted cochlear function, perceptually, patients frequently struggle with sound source localization, temporal perception, and gap detection even in older individuals with normal cochlear function, suggesting dysfunction in central components the auditory pathway, including the auditory cortex (ACtx), the auditory thalamus, and portions of the auditory brainstem and midbrain [3,4,5,6,7,8,9,10] (Fig. 1a). In day-to-day life, central auditory system dysfunction often affects older individuals, causing them to struggle to discriminate speech from a noisy background [7, 11].
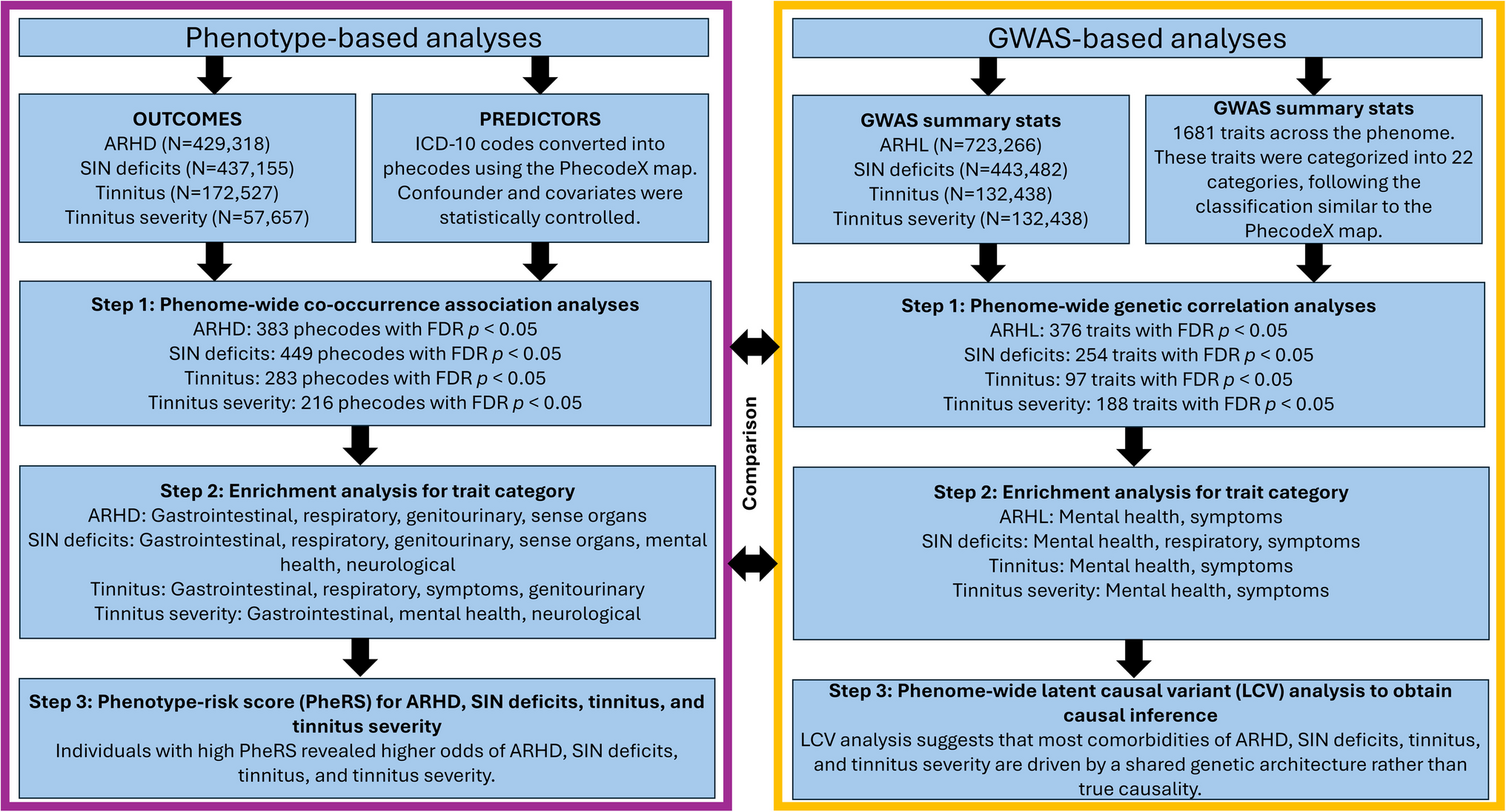
Fig. 1
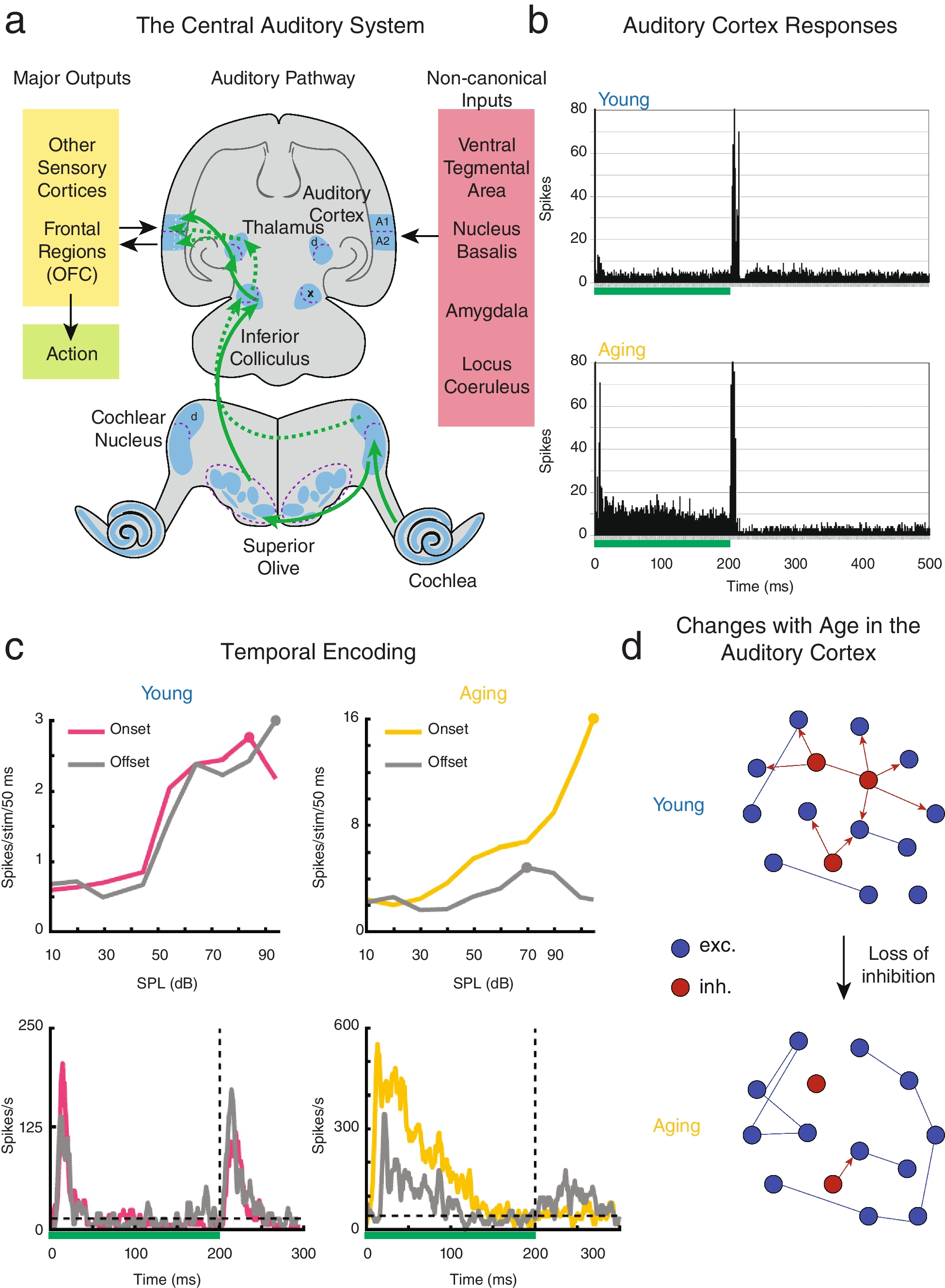
The aged auditory cortex represents auditory stimuli with less specificity. a Schematic of the central auditory system necessary for proper auditory processing. Adapted from Fig. 2.2 of Goodrich & Kanold, [199]. The lemniscal auditory pathway is shown with the solid arrows, and the non-lemniscal pathway is shown with dotted arrows. b Adapted from Fig. 6 of Turner et al., [26]. Collapsed peristimulus time histogram from layer V primary ACtx neurons with complex response profiles from young and aging rats in response to a 200 ms current pulse ranging from 1 to 10 nA (represented by green bar). c Adapted from Fig. 2 of Ramamurthy & Recanzone, [20]. Exemplar ON and OFF intensity rate-level functions (top) and peristimulus time histograms (bottom) from both young and aging non-human primate neurons in response to a 200 ms unfrozen Gaussian noise stimulus (represented by green bar). d Schematic containing the main point made in this figure: as inhibition is lost in aging, the network becomes increasingly homogenous in its stimuli encoding. Circles represent neurons, and lines represent connections. Excitatory neurons are in blue and inhibitory neurons are in red
Cochlear dysfunction is exceedingly common in adult humans [12], likely due to exposure to harsh auditory environments, such as noise pollution, prolonged use of personal listening devices at high volumes, or loud environments. Additionally, independent dysfunction can also be observed throughout the remainder of the ascending auditory pathway in the majority of the aging population, contributing to this auditory dysfunction, although the prevalence of central auditory system dysfunction is difficult to measure in isolation from peripheral auditory system loss [4, 13,14,15,16]. To disambiguate central auditory system dysfunction from peripheral hearing loss, carefully controlled animal studies have been used. To this effect, multiple studies have searched for and found independent effects of aging on ACtx in non-human primates, rats, and mice [17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32]. Studies evaluating the effects of aging on single neurons in the ACtx of the macaque monkey suggest that aging increases both the spontaneous and sound-driven activity of these neurons [18, 19], increasing the effects of noise on auditory processing. These findings significantly advanced the field’s mechanistic understanding of the basis of age-related dysfunction in the ACtx and closely fit the human phenotype of decreased fidelity in auditory perception. To supplement this new knowledge, mouse models were used due to the wide availability of transgenic methods that enable the simultaneous recording of the activity of thousands of neurons in large cohorts of animals raised in controlled housing conditions designed to minimize variations in peripheral hearing experience not easily possible with non-human primates [21,22,23]. Here, we attempt to synthesize literature obtained from humans, non-human primates, rodents, and a variety of other species about aging in the ACtx.
The ACtx is thought to represent sound stimuli using sparse neural networks of excitatory and inhibitory neurons that respond preferentially to specific stimuli composed of numerous variables, such as the frequency, intensity, or location of the sound [33,34,35]. In aging, numerous changes occur in the ACtx, ranging from brain hemisphere-scale changes in the asymmetric processing of spatially distributed sounds [36] to smaller-scale, network-scale changes in the excitation-inhibition balance in local cortical circuits [25]. Over the past decade, a large body of literature has supported the idea that decreased inhibition is responsible for most dysfunctions observed in the aging ACtx [32, 37,38,39,40,41,42] although changes can also be seen throughout the entire auditory pathway [42]. However, recent findings suggest that changes in the ACtx involve both decreased inhibition and altered excitation rather than decreased inhibition alone [21, 22, 24, 25]. Therefore, this review describes the recent advances in our understanding of aging in the ACtx.
Throughout the entire auditory system, auditory stimuli can be primarily characterized by their frequency spectrum, temporal envelope, and binaural differences [43]. Within the ACtx, neurons then integrate this information into complex readouts of multiple components of each auditory stimulus, including its spatial location, amplitude, spectral and temporal components, including as sound onset or offset leading to high stimulus selectivity [33,34,35, 43,44,45,46,47]. Furthermore, sound features are also encoded by neuronal ensembles in ACtx [21, 45, 46, 48, 49] and neuronal responses are modulated by arousal and behavioral state, such that neural and ensemble responses vary between passive tone presentation and during behavioral tasks [22, 47,48,49,50,51,52,53]. Sex has also been shown to affect the activity of neural populations and circuits in the ACtx, especially during aging [21, 22, 24]. Given the processing complexity in the brain, even slight circuit and functional changes associated with aging can have a dramatic impact on cognitive abilities, including those in the sensory systems. However, the detailed circuit and processing changes in the ACtx with aging were not well-studied until recently. The cerebral cortex demonstrates a tremendous capacity for plasticity, and an expanding body of research highlights promising strategies, such as auditory training and vagal nerve stimulation, to restore or preserve auditory cortical function [23, 54,55,56,57,58]. Thus, this review will focus on the changes in the primary ACtx that disrupt the spectral and temporal representations of sound before concentrating on the effects of sex and plasticity in the aging ACtx. While this work focuses on the primary ACtx, all ACtx references can be assumed to refer to the primary ACtx unless otherwise stated. Available research about the secondary ACtx is referenced when available, but differences in aging between primary and higher-order areas remain a central open question.
Decreased Functional Response Selectivity in the Aging Auditory CortexThe aged ACtx fundamentally differs from the young ACtx in both structure and function. Thus, the aged ACtx represents stimuli differently and often with less fidelity than the young ACtx. Research conducted in humans has revealed distinct cortical activity patterns in young and older adults when listening to auditory stimuli. When an individual listens to auditory stimuli, the ACtx performs a type of pattern recognition, allowing for proper speech recognition [59, 60]. However, sustained neural activity resulting from this speech recognition pattern appears to be lost in the aged human ACtx when measured using magnetoencephalography (MEG) [61]. Accordingly, speech has been shown to recruit more neurons in the ACtx of older individuals as compared to young individuals [62, 63], suggesting that decreased auditory processing abilities, which limit speech comprehension, correlate with an overrepresentation in, and thus less specific, neural responses [64]. As such, older listeners often place excessive emphasis on the context of speech to aid in word recognition, which can lead to a misperception of the actual spoken word, a phenomenon termed false hearing [65]. This, too, has been hypothesized to be a result of decreased inhibition of high-probability responses, leading to the misperception of speech [66].
Moreover, multiple studies in humans have found that a decrease in inhibition, as measured by magnetic resonance (MR) spectroscopy, in the ACtx correlates with impaired speech-in-noise comprehension [38, 39, 67]. However, due to the inherent limitations of working with human subjects, these experiments lacked many finer observations made possible by invasive experiments in animal models. Thus, studies using non-human primates nicely complement human studies by allowing for direct, temporally precise measurements of neural activity in the ACtx. Such studies have revealed that auditory stimuli cause a higher spike rate of individual excitatory neurons in the aged ACtx, in addition to an increase in spontaneous activity present in aged neurons [17,18,19], perhaps partially due to a compensatory loss of inhibition in the ACtx to make up for decreased peripheral excitation in the case of accompanying peripheral hearing loss [26, 68]. Thus, the increase in cortical activity, concomitant with a decrease in functional response specificity to any given sound, suggests a fundamental change in the underlying circuitry of the ACtx throughout one’s lifespan.
With the advances in transgenic models, the relative ease of controlled housing, and shorter lifespan, single-neuron recordings and circuit-level in vivo calcium imaging in rodents have proven invaluable for studies on the individual neuron and circuit levels. Early studies that recorded single-neuron activity in response to a GABAA receptor antagonist indicated that local inhibition plays a crucial role in maintaining sparse tuning in the ACtx [69, 70], and subsequent molecular and slice-physiology studies found that aging disrupts levels of inhibition, leading to broader tuning throughout the ACtx [40,
Comments (0)