
Remember me

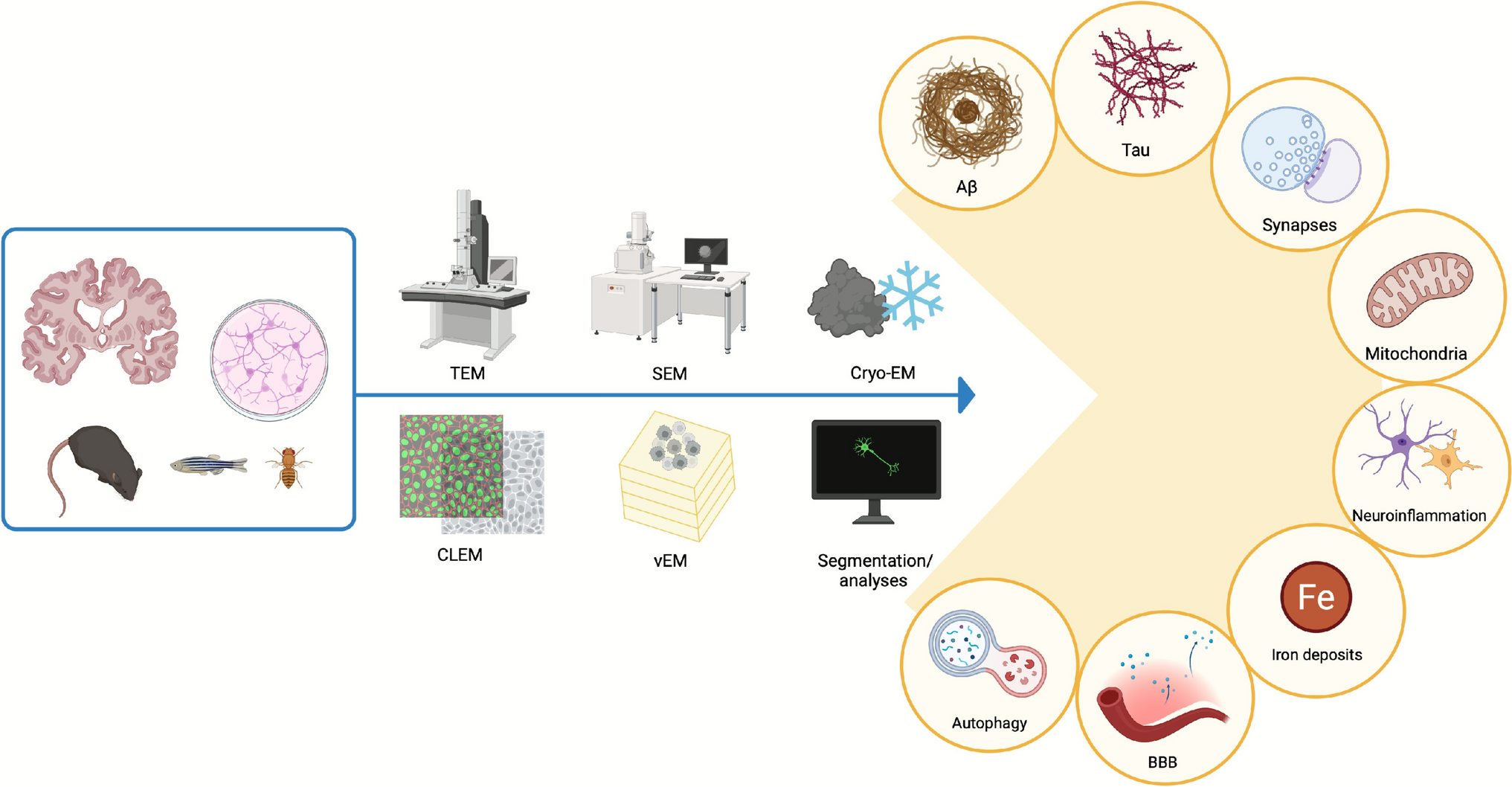


The amyloid cascade hypothesis of Alzheimer disease suggests that a pathological event is the buildup of Aβ peptides within the brain (Selkoe & Hardy 2016). This theory is largely supported by neuropathological findings and human genetic evidence, particularly from familial cases of AD (FAD), while it is less applicable to sporadic AD (SAD) (Frisoni et al. 2022). Moreover, new Aβ-targeting drugs, such as Lecanemab, were successful in reducing amyloid burden, but they had moderate effect in slowing cognitive decline in early AD, in some cases at the expense of severe adverse effects, in clinical trials (Van Dyck et al. 2023).
In this context, a more accurate knowledge of the timing of Aβ deposition, of the behavior of different Aβ fragments and of their interplay with other AD hallmarks (tau pathology, neuroinflammation, synaptic and mitochondrial dysfunction etc.) is critical.
Senile plaques were first described in 1906 by Alois Alzheimer, and in full detail by his mentee Gaetano Perusini in 1909 (Rosso and Chu 2021), but only with the spread of EM, starting from the ‘60 s, their composition was clarified. Terry et al. (1964) (Terry et al. 1964) described for the first time the fibrillary core of plaques in cerebral biopsies of three cases of early onset AD. The central core was described as a “stellate mass of interwoven fibers, each 70 to 90 Å wide”; fibrils were grouped into bundles, generally oriented toward the center of the plaque and devoid of membranes. It was clear that central fibrils and bundles were extracellular, while those in the plaque periphery were found in close relationship to nuclei and organelles. Meanwhile, Shirahama and Cohen were able to describe the general structure of all amyloid fibrils: they are made of 5 parallel protofibrils (25–35 Å), which are in turn made of 2 or 3 subprotofibrils helically arranged (10–15 Å) (Shirahama and Cohen 1967). In 1980, these structures were also described at the ultrastructural level in brain biopsies from AD patients (Narang 1980).
In 1984, amyloid twisted β-pleated sheet fibrils were then purified from AD brain (Glenner and Wong 1984) and their amino acid sequence was found to have no homology with other known proteins. In 1985, immunogold EM labeling was used to demonstrate in vitro fibril formation from two synthetic peptides homologous to β-amyloid fragments (Castaño et al. 1986) and in 1987 similar results were obtained through x-ray diffraction and EM (Kirschner et al. 1987), thus allowing the identification of the specific sequence responsible for AD-like fibril formation. Finally, in 1989 Halverson et al. showed that actually the sequence 34–42 is responsible for the formation of stable, insoluble β-structures (Halverson et al. 1990).
Recent AdvancesSince the 2010s, advances in both conventional EM and cryo-EM have significantly enhanced our understanding of Aβ ultrastructure (view Table 2 for further studies). Notably, cryo-EM has enabled near-atomic level visualization of Aβ fibrils. In 2017, Gremer and colleagues reported the structure of recombinant Aβ42 fibril, revealing two twisted protofilaments, by employing cryo-EM at 4.0 Å resolution in combination with solid-state nuclear magnetic resonance techniques. They also resolved the Aβ42 backbone of 42 residues, including the entire N-terminus (Gremer et al. 2017).
This structure was later refined and complemented by subsequent studies (Ghosh et al. 2021), which uncovered further conformational polymorphisms across fibrils purified from patients’ meninges or cortex. Moreover, Yang et al. (Yang et al. 2022) reported that Aβ42 extracted from human brain tissue forms two types of filaments. Type I was predominant in SAD patients, whereas Type II was mainly observed in familial AD (FAD) cases and represented the dominant form in AppNL-F knock-in mice (Table 2).
These results underscore the high heterogeneity of amyloid fibrils and aggregates; indeed amyloid deposition was demonstrated to be different among diverse tissues (Yang et al. 2023b, a), in various mice models (Zielinski et al. 2023) and in recombinant Aβ fibrils that were formed in vitro (Kamalaldinezabadi et al. 2024; Zielinski et al. 2025) (Table 2).
Table 2 Cryo-EM contribution to the understanding of Aβ structure: this table summarizes key studies describing Aβ conformation, with the use of Cryo-EM alone or in combination with other techniquesThese studies further emphasize that investigating structural variations in Aβ fibrils may help future investigations linking Aβ profiling with severity, progression rate or clinical manifestations in AD (Yang et al. 2022). Understanding these variations is therefore crucial for developing inhibitors and imaging agents with both therapeutic and diagnostic potential (Ghosh et al. 2021).
Another striking result of the most recent ultrastructural studies is that the Arctic (E22G) mutation in amyloid precursor protein (APP) enhances Aβ40 fibril accumulation and cross-propagation in FAD, with higher Aβ40 deposition in plaque core compared to SAD (Tehrani et al. 2024).
Despite these observations, the in situ structure of amyloid in human brain remained elusive and was elucidated by Gilbert et al. only in 2024 (Gilbert et al. 2024) (Fig. 1A). Using cryo-correlative light-electron microscopy (CLEM) and cryo-ET, it was shown that Aβ plaques are composed of a heterogeneous mixture of fibrils—including branched forms—and protofilaments, organized in parallel arrays and lattice-like architectures. In addition, the plaques were found to be interwoven with various membrane-bound subcellular compartments, such as extracellular vesicles (EVs), lipid droplets and fragmented membranes.
Fig.1
Representative EM novel findings in AD. Images are adapted from previously cited, open-source papers
Beyond allowing its structural characterization, EM has been pivotal in localizing Aβ at subcellular resolution. Indeed, recent investigations on thin sections of AD cerebral cortex found that intracellular Aβ α-sheet structures are intracellularly associated to lipofuscin, although pathobiological implication of this association are unknown. This raises new hypotheses regarding the formation of toxic oligomeric intermediates (Serwer et al. 2022) and shifting the focus from fibrils to soluble assemblies in disease initiation.
Besides these outstanding results, advanced immunogold EM techniques have shown that oligomeric Aβ species accumulate within synaptic terminals in a mouse model of AD, while array tomography has allowed the identification of different oligomeric species present in these subcellular structures (Pickett et al. 2016). Indeed, Aβ oligomers are known to interact with the neuronal membranes and induce perforations that cause an influx of calcium ions and enhance the release of synaptic vesicles, leading to delayed synaptic failure by vesicle depletion, as proved by TEM (Parodi et al. 2010).
Furthermore, the combination of different techniques, such as surface plasmon resonance and immunogold labeling, followed by direct analysis of the sensor-chip surface by SEM, has allowed to observe the lateral binding of Apolipoprotein E along the amyloid fibrils (Islam et al. 2019), thus opening the avenue for identifying novel potentially pathogenic interactions of Aβ.
In in vitro studies, the effect of Aβ on different cell types has also been elucidated via EM. For instance, Aβ was able to induce excessive mitochondrial fission in endothelial cells, leading to apoptosis (Zhang et al. 2025). Similarly, Aβ exposure was shown to induce pyroptotic features in cultured astrocytes (Hong et al. 2023).
Interestingly, recent EM-based investigations have also extended the study of Aβ beyond the traditional CNS tissue. Indeed, one challenge in neurodegenerative diseases is to identify peripheral biomarkers as minimally invasive diagnostic tools to be exploited at early or even preclinical stages of the disease. Notably, Aβ aggregates have been identified by immunogold EM in the cytoplasm of supranuclear fiber cells of the lenses in both human patients and murine models of AD and Down syndrome (Moncaster et al. 2010). In the same context, SEM and immunogold labeling have allowed to observe Aβ aggregates in periodontal biofilms (Kanagasingam et al. 2022) and in circulating neuron-derived extracellular EVs (Eitan et al. 2023).
Finally, EM has also been a useful tool to characterize alternative models of AD involving Aβ accumulation. For instance, in both APPL-RNAi and transgenic Drosophila melanogaster lines expressing eye tissue-specific human Aβ, eye degeneration was reported through SEM (Singh et al. 2017); while in a SAD-zebrafish model, TEM has highlighted the presence of higher order insoluble amyloid fibrils with twists (Dhiman et al. 2025).
Taken together, the evidence presented here highlights how EM has been fundamental in the initial characterization of senile plaques and how, in recent years, its progressive refinement has been pivotal in elucidating the ultrastructural complexity and localization of Aβ aggregates, as well as in characterizing novel models of Alzheimer disease—thereby significantly advancing our understanding of its pathogenesis.
TAUBackgroundThe microtubule associated protein tau has been identified as the main component of the insoluble aggregates that characterize a number of neurodegenerative diseases collectively known as tauopathies (Kovacs et al. 2022). In AD, the commonest type of tauopathy, abnormally phosphorylated tau (pTau) accumulates in form of NFTs, dystrophic neurites and neuropil threads. The importance of tau dysfunction in AD is underscored by the fact that the degree of cognitive impairment and dementia correlates strictly with the amount of tau burden in AD brain (Creekmore et al. 2024b). Given the failure of all currently available pharmaceutical trials aimed at reducing the amount of Aβ in AD patients, elucidating the role and mechanism of tau accumulation becomes essential for developing alternative and effective therapeutic strategies; studies of pTau at the ultrastructural level are therefore of the outmost importance.
In AD, tau undergoes abnormal phosphorylation and as a consequence detaches from microtubules, aggregates into paired helical filaments (PHFs) and straight filaments (SFs) and accumulates in form of NFTs (Ikeda et al. 1992). The twisted nature of the aggregates was first shown at the ultrastructural level in 1963 by Kidd (Kidd 1963). Early studies using conventional TEM detected ~ 15 nm thick straight and twisted tubules within neurons, degenerating neurites and in the extracellular “ghost” tangles (Crowther 2021). Subsequent studies using immunogold techniques at the ultrastructural level helped identifying tau as a protein associated to microtubules within normal tissue (Migheli et al. 1988), as well as the main component of PHFs and SFs in AD brain in its abnormally phosphorylated form (Crowther and Goedert 2000). Soon it was shown by immunogold EM that pTau localizes not only to NFTs, but also to ribosomal and periribosomal granular regions of neurons, suggesting a role of pTau in early impairment of the translational machinery (Papasozomenos 1989).
Recent AdvancesUltrastructural evidence of the association to ribosomes has been recently confirmed with a panel of antibodies to multiple pTau epitopes, which revealed that pTau association in granular pretangles and perinuclear zones precedes the appearance of more mature fibrils, thus implying mislocalization of tau as an early sign of neuronal dysfunction (Price et al. 2025). By using advanced techniques such as quantum dot labeling for correlating LM and EM, it was shown that LM-defined pretangles are actually tau-positive SFs appearing before classical NFTs become recognizable, and that the AD pretangles have a distinct appearance from those of other tauopathies (Tatsumi et al. 2014).
Recently, the development of cryo-EM techniques at a near-atomic resolution has provided a tremendous advancement in clarifying the formation and the role of pTau aggregates in AD (further details are available in Table 3). By mapping PHFs and SFs at 3.4–3.5 Angstrom resolution, Fitzpatrick et al. (Fitzpatrick et al. 2017) first showed that filament cores are made of two identical protofilaments comprising residues 306–378 of tau protein, which fold into a cross-β/β-helix structure, and that PHFs and SFs differ in their protofilament packing, definitely demonstrating ultrastructural polymorphism of the tau aggregates. Subsequent cryo-EM studies have further mapped the tau fold, showing that it differs from those described in other tauopathies, and identifying an AD core region of 79 aminoacids with five β-strands, which gives rise to two antiparallel β-sheets plus a β-helix region that also serves as a target for high-affinitiy PET ligands (Shi et al. 2021). Interestingly, cryo-EM studies in Down syndrome (DS), a condition with increased risk of developing early onset AD, have revealed PHFs and SFs made of C-shaped protofilaments with the same cross-β/β-helix structure of AD, underscoring the ultrastructural similarity at the molecular level of tau pathology across AD-related diseases (Ghosh et al. 2024).
Table 3 Cryo-EM contribution to the understanding of tau structure and filament formation: this table summarizes key structural findings regarding tau filaments with the use of Cryo-EM alone or in combination with other techniquesMoreover, templated seeding studies in SH-SY5Y cells seeded with extracts derived from AD and other tauopathies have shown by cryo-EM some degree of templating, suggesting that tau filaments in vitro resemble disease-specific conformations (Tarutani et al. 2023), thus supporting a prion-like model of propagation of toxic tau (Dujardin and Hyman 2019). The latter hypothesis is also supported by findings of an association of assembled tau with EVs (Ruan et al. 2021). A better understanding of the molecular mechanism underlying EV-mediated secretion of tau has been provided by recent cryo-ET and single-particle cryo-EM studies, that revealed truncated tau contained within EVs enriched in endo-lysosomal proteins, along with interactions between tau filaments and molecules that anchor them to the EV limiting membrane, suggesting a process of selective packaging (Fowler et al. 2025).
Attempts to assembly in vitro recombinant tau into filaments that are structurally identical to PHFs of AD have been successfully shown by cryo-EM analysis (Lövestam et al. 2022). These studies are of the greatest importance, as in vitro reconstructed tau filaments may clarify the relationship between ultrastructure, aggregation/folding kinetics, phosphorylation dynamics, thus opening new avenues for developing chemical compounds which may interfere with phosphorylation and fibril formation (Huseby and Kuret 2016).
Another powerful application of ultrastructural EM techniques has been directed to understanding the assembly of soluble tau oligomers purified from AD brains. Negative-staining TEM and atomic force microscopy (AFM) showed that pTau assembles into misfolded highly heterogeneous oligomers. The bigger oligomers with a diameter of 10–20 nm were observed in Cryo-EM (with a 2.5–4 Å resolution), which revealed ring-like structures with two thin layers separated by approximately 1 nm; this structure could enhance their interactions with cellular membranes and proteins (Dasari et al. 2025).
Comparative ultrastructural studies have allowed to analyze and classify tau polymorphisms across the various tauopathies: cryo-EM and NS-TEM analyses can differentiate AD from Pick disease, progressive supranuclear palsy and corticobasal degeneration, based on fold length, β-helix structure and protofilament organization, supporting the strain propagation model of distinct toxic tau conformations (Creekmore et al. 2021, 2024b; Goedert 2021; Lövestam et al. 2022).
While TEM and cryo-EM have generated an impressive amount of data regarding the formation and assembly of pTau subcellular structures in AD, SEM techniques have been fundamental for elucidating the three-dimensional structure of NFTs, revealing tau positive filamentous assemblies in situ with nanometer scale resolution. In a seminal paper using freeze cracked human autopsy tissue, both PHFs and SFs were readily distinguished, the first with a diameter of 28 nm and periodic constrictions of 100 nm, the second with a diameter of 20–25 nm. In addition, atypical variants were observed, supporting a protofilament model of PHFs (Itoh et al. 1997).
On the other hand, scanning TEM (STEM) has been employed to directly determine the mass-per-length (MPL) of individual tau fibrils isolated from AD brains, showing that a single polypeptide can give rise to both polymorphic fibrils and diverse globular oligomers simultaneously (Goldsbury et al. 2011). Using cryo-ET, combined with cryo-FIB-SEM lift-out to prepare tissue lamellae, tau inclusions were observed forming parallel arrays of unbranched filaments. Subtomogram averaging of 136 tau filaments within a single tomogram allowed determination of the polypeptide backbone conformation and the filament polarity of paired helical filaments (PHFs) in tissue (Gilbert et al. 2024) (Fig. 1B).
Using SEM, energy dispersive spectroscopy (EDS) and electron energy loss spectroscopy (EELS), pTau deposits were shown to colocalize with iron and zinc deposits in AD brain, suggesting a link between tau pathology and oxidative metal deposition (Madsen et al. 2020).
Finally, the recent advances in vCLEM and the availability of large datasets have allowed the identification of an unusual localization of pTau, i.e. inside spine-like protrusions at the axon initial segment. These structures could be prone to detachment and, therefore, might be involved in the formation of exosomes—a process implicated in the dissemination of pTau in AD (Han et al. 2023).
In summary, SEM techniques have provided useful data to elucidate not only the morphology of tau aggregates (bundle dimensions, filament diameter, periodicity), but also the spatial localization within neurons and the pathological microenvironment of NFTs, thus complementing the traditional investigation of tau with TEM and cryo-EM.
As a final word, data from ultrastructural studies have also been exploited to develop mathematical models of tau propagation across brain regions. Using data from imaging and ultrastructural staging studies, these theoretical models can predict region-specific spreading patterns of tau, not only by diffusion, but also by axonal transport, consistent with the clinical progression (Tora et al. 2025).
SynapsesBackgroundSynaptic degeneration is already evident in AD early-stages, preceding neuronal loss, and is one of the earliest and most predictive features of AD (Meftah and Gan 2023). Accordingly, synaptic loss functionally correlates with cognitive impairment in AD (de Wilde et al. 2016a, 2016b).
Axons and dendrites have long been recognized as key components in AD pathology. Terry et al. (1964) observed that, in the periphery of plaques, these structures appeared enlarged and contained abnormal neurofilaments and, occasionally, dense bodies (Terry et al. 1964). Similarly, Krigman in 1965 and later Gonatas (Gonatas et al. 1967) reported enlarged presynaptic endings that also contained dense bodies.
This concept gained quantitative support over the following decades. In 1990, DeKosky et al. demonstrated an important reduction in synapse density in the frontal cortex of AD patients. Accordingly, there was a significant correlation between synapse count and Mini-Mental State examination scores (DeKosky and Scheff 1990). The synaptic degeneration was further characterized by immunogold EM labeling, thus reinforcing the hypothesis of progressive synaptic pathology in AD neocortex (Masliah et al. 1991).
Taken together, these foundational studies laid the groundwork for our current understanding of AD as a synaptopathy and established EM as a crucial tool for exploring the disease’s earliest microstructural changes.
Recent AdvancesIn recent years, the advent of advanced vEM techniques, such as FIB-SEM, has allowed the micro-anatomical characterization of the neuropil of brain regions involved in AD, revealing region-specific synaptic loss. It was thus shown that while transentorhinal cortex thickness is reduced in AD, leading to a general loss of overall synapse number, the synaptic density is maintained (Domínguez-Álvaro et al. 2018). In this area, asymmetric synapses, that are classified as excitatory based on their post-synaptic density (Peters and Palay 1996), were more fragmented than in controls (Domínguez-Álvaro et al. 2019). Likewise, when the same technique was applied to the CA1 area of the hippocampus, early cases of AD did not display changes in synapse density, nor in excitatory/inhibitory synapse ratio, although there was already evidence of cortical atrophy and of changes in the distribution of postsynaptic targets and synaptic shapes. On the other hand, late AD cases suffered a decrease in synaptic density and morphological alterations of the remaining synapses (Montero-Crespo et al.
Comments (0)