We reconstructed the functional connectivity gradients of the pons based on its connectivity with the cerebral and cerebellar cortices to uncover principles governing its functional organization. We found that the primary functional gradient of the pons was organized in a rostral-to-caudal and medial-to-lateral fashion that, when projected onto the cerebral and cerebellar cortices, formed gradients that corresponded closely with the unimodal-to-transmodal organization seen in prior work (Guell et al. 2018; Margulies et al. 2016). Rostral and medial areas of the pons were preferentially connected to default mode and frontoparietal network areas, whereas caudal pons aligned with somatomotor, dorsal and ventral attention network areas of the cerebral and cerebellar cortices. Our results align closely with previous gradient work, but residuals indicated subtle but systematic differences localized in specific cortical and cerebellar areas suggesting overlapping network represents within particular areas of the pons. Together, these findings provide supporting evidence that the pons has a complex functional organization and may serve as an integrative hub in the downstream corticocerebellar pathway.
A rostral-caudal and medial-lateral functional connectivity gradient in the pons is remarkably consistent with anatomical connectivity work in non-human primates (Schmahmann et al. 2004; Schmahmann and Pandya 1997b) and our recent dMRI tractography work in humans (Rousseau et al. 2022, 2025). Specifically, motor projections to the pons have been shown to terminate preferentially in the caudal half of the pons (Schmahmann et al. 2004), which is also the origin of pontocerebellar fibres projecting to cerebellar motor areas (Brodal 1979; Rousseau et al. 2022). Prefrontal cerebrocortical areas target the medial pons (Schmahmann and Pandya 1997a). Medial prefrontal cortex targets rostral pons whereas dorsolateral prefrontal projections are distributed along its rostral-caudal extent (Schmahmann and Pandya 1997a). Different areas of the parietal cortex also show differential patterns of projection to the lateral pons, with the inferior parietal lobule projecting rostrally and the superior parietal lobule projecting more diffusely along the rostro-caudal extent (Schmahmann and Pandya 1989). These anatomical findings are mirrored in our primary gradient, which shows sensorimotor areas clearly localized to the caudal pons and a more complex, and distributed pattern for non-sensorimotor regions. Specifically, medial prefrontal areas and the inferior parietal lobule – both nodes in the default mode network – correspond to the medial and rostral portions of the pons in our gradient. Conversely, the superior parietal lobule and portions of dorsolateral prefrontal cortex show a wider distribution of connectivity with the caudal two thirds of the pons. Broadly, our findings correspond with organizational patterns demonstrated in the anatomical literature.
While our findings align with certain aspects of known anatomical connectivity, they diverge in ways that highlight the complexity of comparing structural and functional connectivity. For instance, detailed work in macaques has demonstrated that temporal lobe corticopontine projections arise predominantly from the upper bank of the superior temporal sulcus and the superior temporal gyrus (Schmahmann and Pandya 1991). However, in our gradient, we found that the rostral pons maps onto large areas of middle and inferior temporal gyri. Without excluding the possibility that the anatomical connectivity is different in humans, this is more likely to reflect indirect connections mediated at the level of the cerebral cortex (Damoiseaux and Greicius 2009). These temporal regions are highly interconnected with other cortical areas with known projections to the pons in non-human primates (e.g., inferior parietal lobule). It follows that the functional connectivity of the pons, and by extension our gradient, is shaped by a combination of direct and indirect cortical inputs. In addition, though we excluded pons-pons connectivity from our analysis we cannot rule out potential indirect contributions that could be mediated by pontine interneurons (Mihailoff et al. 1992) and/or projections from the deep cerebellar nuclei (Schwarz and Thier 1999).
Our findings also differ from those of Margulies (2016) and Guell et al. (2018) in a manner that may suggest a possible integrative function in the pons. In our residual analysis, when comparing our gradient projections with the primary gradient of Margulies (2016), we found strong negative residuals in visual cortex. This finding suggests that (relative to the canonical gradient presented by Margulies (2016) visual areas in our gradient projection are situated towards the middle of the gradient, whereas in Margulies (2016) these same areas clearly correspond to the unimodal end of their primary gradient. The anatomical tract-tracing work provides compelling evidence that ventral stream visual areas project along the rostro-caudal extent of the pons (Schmahmann and Pandya 1993). With our dataset, connectivity from visual areas to the pons do not appear to be spatially specific, in line with these anatomical findings. This results in the projection to this area sampling values from both ends of the pontine gradient, leading to more intermediate values. By extension, this also implies that visual inputs to the pons are in proximity to both motor inputs and inputs from associative cortical areas, thus providing the spatial proximity necessary for integration (Schwarz and Thier 1999). In our residual analyses, we also found strong positive residuals in the inferior frontal gyrus, middle temporal gyrus, and regions of the cerebellar cortex that are recruited during language production tasks (King et al. 2019). This pattern suggests that, in our gradient projection, these language related regions are shifted towards the unimodal end of the functional hierarchy relative to the cerebrocortical and cerebellar cortical gradients of Margulies (2016) and Guell et al. (2018). It may imply that at the level of the pons, language and sensorimotor channels may be more integrated or overlapping than in the cerebral or cerebellar cortices.
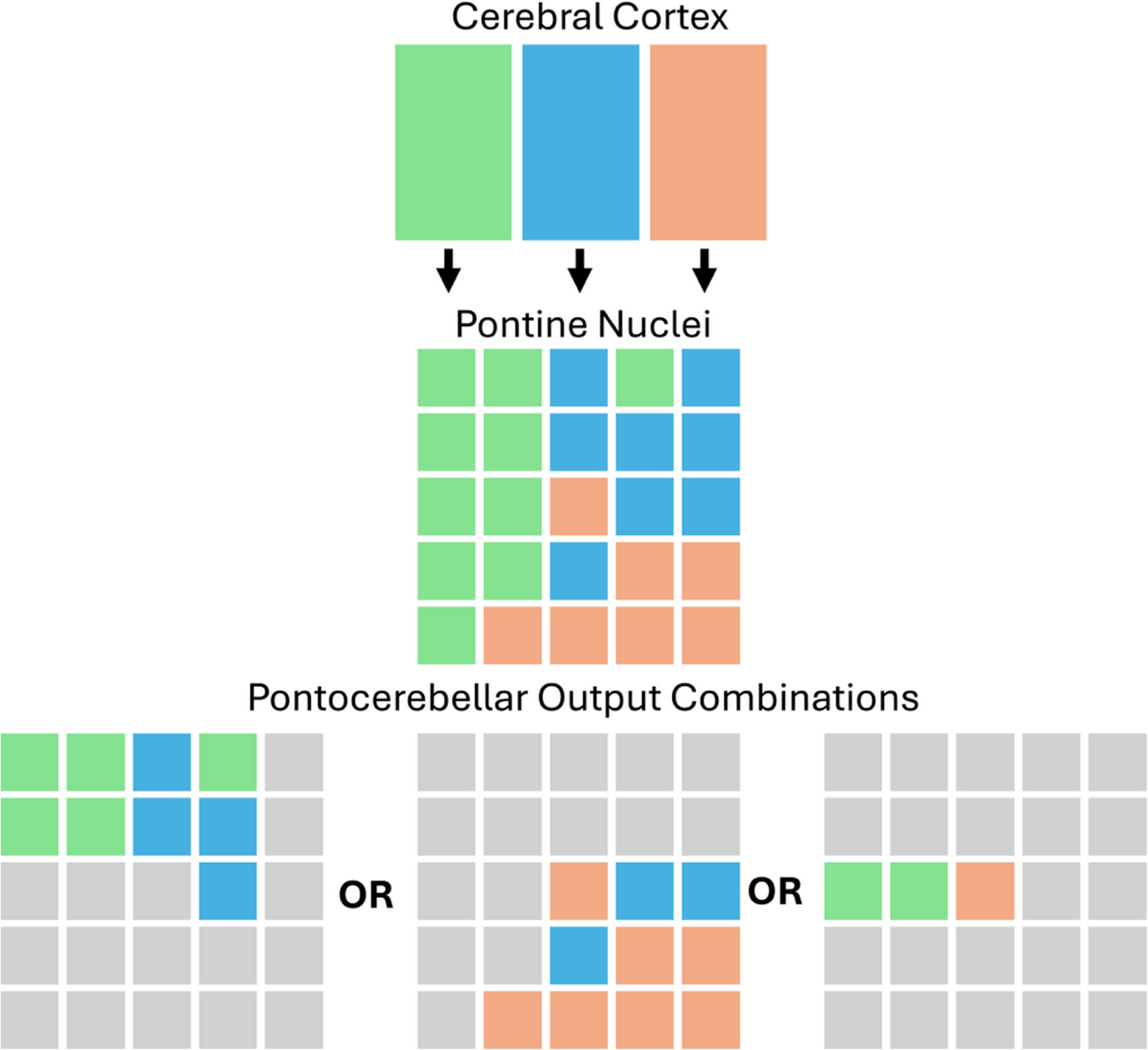
The anatomical tracing work shows that corticopontine terminations from different cerebrocortical areas are interdigitated with one another, bringing inputs from different cerebrocortical areas (e.g., visual and motor) in close proximity to one another. Schwarz and Thier (1999) propose that this arrangement complements filter-like characteristics of pontocerebellar neurons to allow the pons to integrate signals from different regions before projection to the cerebellum as mossy fibres. They suggest that depending on context or specific environmental demands, different combinations of corticopontine inputs can be integrated and relayed to the cerebellum (see Fig. 1). Our findings align with and support portions of this theory by demonstrating the convergence of different functionally relevant inputs to overlapping zones within the pons. Thus, our results provide evidence that visual information may be integrated with motor related signals (e.g., for visually guided reaching) or coupled with non-motor signals (e.g., for non-verbal working memory) at the level of the pons. Similarly, language-related inputs may be integrated with sensorimotor information or with signals from multimodal association areas depending on contextual demands.
The fact that cortical, pontine, and cerebellar nodes of the cortico-ponto-cerebellar pathway appear to be organized in a similar hierarchical fashion may be interpreted as the cortex imprinting its functional architecture on downstream areas. Alternatively there may be more of a reciprocal relationship such that the pons and cerebellum may together influence integration at the level of the cortex. Developmentally, the spatial organization of pontine neurons is established early, and these neurons provide cues that guide the termination patterns of corticospinal axons (Kratochwil et al. 2017). This suggests that the pons actively reshapes the configuration of cerebrocortical inputs, rather than merely reflecting their organization. The gradients observed in the present study likely reflect a combination of macroscale organizational principles, possibly reflecting the anatomical configuration of these pathways, and influences of experience and cerebellar learning. Schwarz and Their (1999) emphasize the role of projections from the deep cerebellar nuclei to the pons in shaping the information it relays to the cerebellum. Within this framework output from cerebellar nuclei – which is shaped by cerebellar models and learning – plays a role in sculpting the functional organization of the pons. It follows that experiences and pathological factors may influence the organization of pontine gradients, highlighting an important direction for future work.
While the present findings provide a biologically plausible characterization of pontine functional organization, they must also be interpreted within the context of brainstem specific methodological limitations. We cannot rule out that the observed patterns of convergence of functionally distinct inputs in the pons are influenced by the small size of this structure and the level of spatial resolution of the dataset. We also note that relative to prior gradient work (e.g., Guell et al. 2020; Katsumi et al. 2023; Margulies et al. 2016) with the same or similar sample size, the cerebral cortex and cerebellar cortical projection gradients presented here exhibit more “noise”. This can be appreciated as a relative decrease in spatial smoothness across the gradient projections. Even with this large sample of high-quality fMRI data used in the present study, we still face important limitations related to low SNR in the pons that represent a barrier to more granular investigations of its functional topography. For instance, while there may be important information within the other gradients, the potential influence of noise and prominent laterality effects led us to focus our interpretation on the first gradient, which we are confident reflects a robust and meaningful pattern. The brainstem represents a worst-case scenario for functional MRI: it is compact in size with densely packed and functionally distinct nuclei, there is poor differentiation of gray matter nuclei from surrounding white matter at the resolutions possible with in-vivo MRI, and there is both elevated physiological noise and magnetic susceptibility artefacts (Sclocco et al. 2018). Group registration may also lead to misalignment of smaller brainstem nuclei, which serves to further reduce the effective resolution of fMRI (Sclocco et al. 2018). Some of these limitations are partially mitigated by averaging a large number of high qualities rsfMRI datasets, as was also done in the present study. However, this approach may also affect the level of granularity with which we can investigate the pons.
Moving forward, replication of the present findings at higher spatial resolutions using acquisitions more tailored to the brainstem is needed. Improved brainstem data quality would also allow for the extraction of meaningful gradients at the individual participant level. This represents an important goal for future work, as it would enable an examination of individual variability and different experiential or pathological factors shaping pontine gradients. Future fMRI studies of the pons would benefit from the collection of physiological recordings, use of models of the hemodynamic response function tailored to the brainstem, probabilistic atlases of brainstem nuclei that can improve inter-subject registration, and the use of high-field (e.g., 7 T) MRI data (Groot et al. 2024; Sclocco et al. 2018). Mohamed and colleagues (2024), for instance, developed a specific protocol for brainstem fMRI with 3 T and demonstrated significant improvements in SNR compared to an HCP style protocol. Improved brainstem data quality will lend itself to understanding the spatial distribution of pontine activation in task-based paradigms (e.g., similar to the approach of King et al. (2019) for the cerebellum), allow for the further delineation of pontine resting connectivity gradients and help clarify how these patterns vary across individuals.




Comments (0)