
Remember me

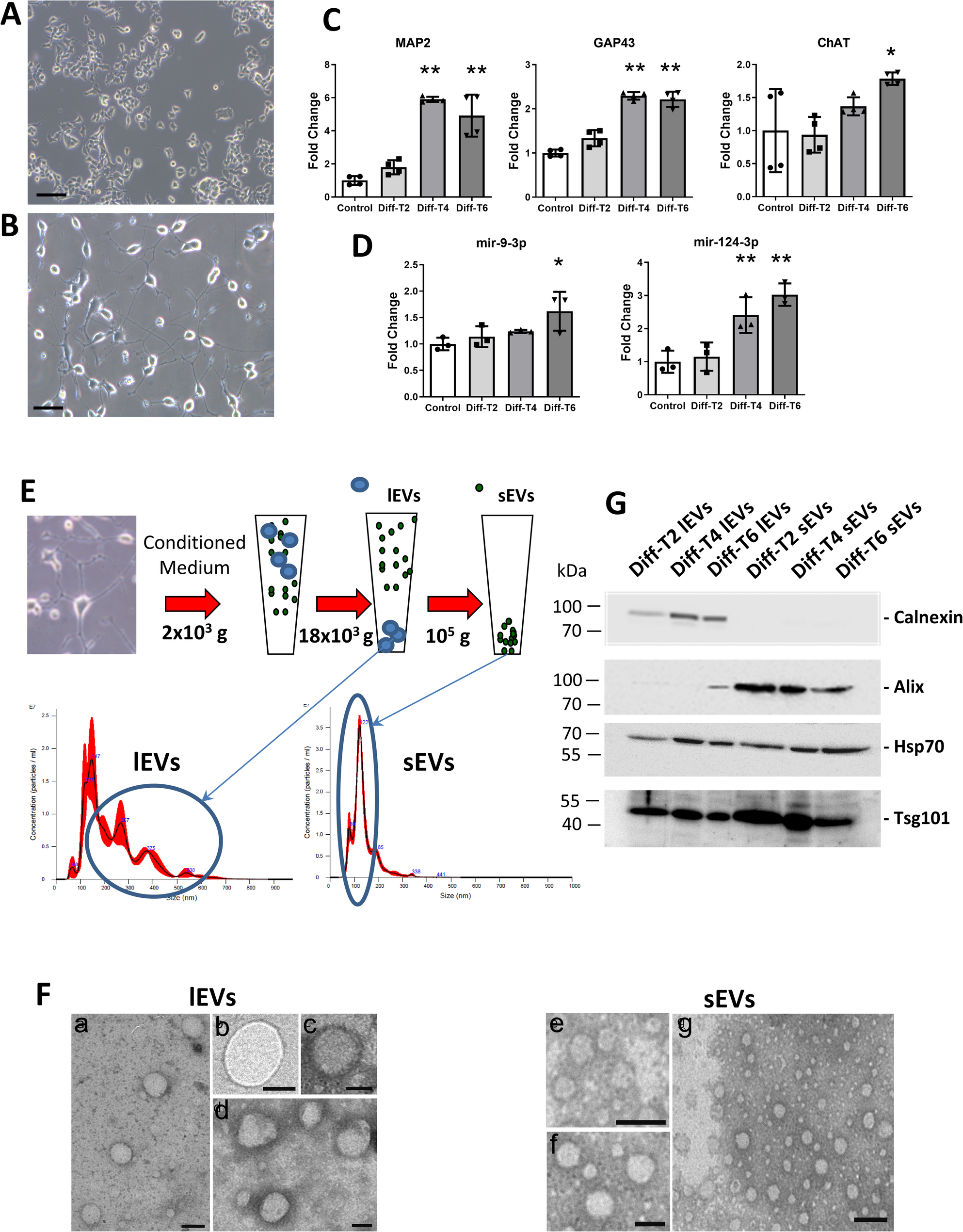

The NSC-34 cells express many of the morphological and physiological properties of motor neurons, including extension of processes, formation of contacts with cultured myotubes, synthesis and storage of acetylcholine, support of action potentials, induction of myotube twitching, and expression of neurofilament proteins [35].
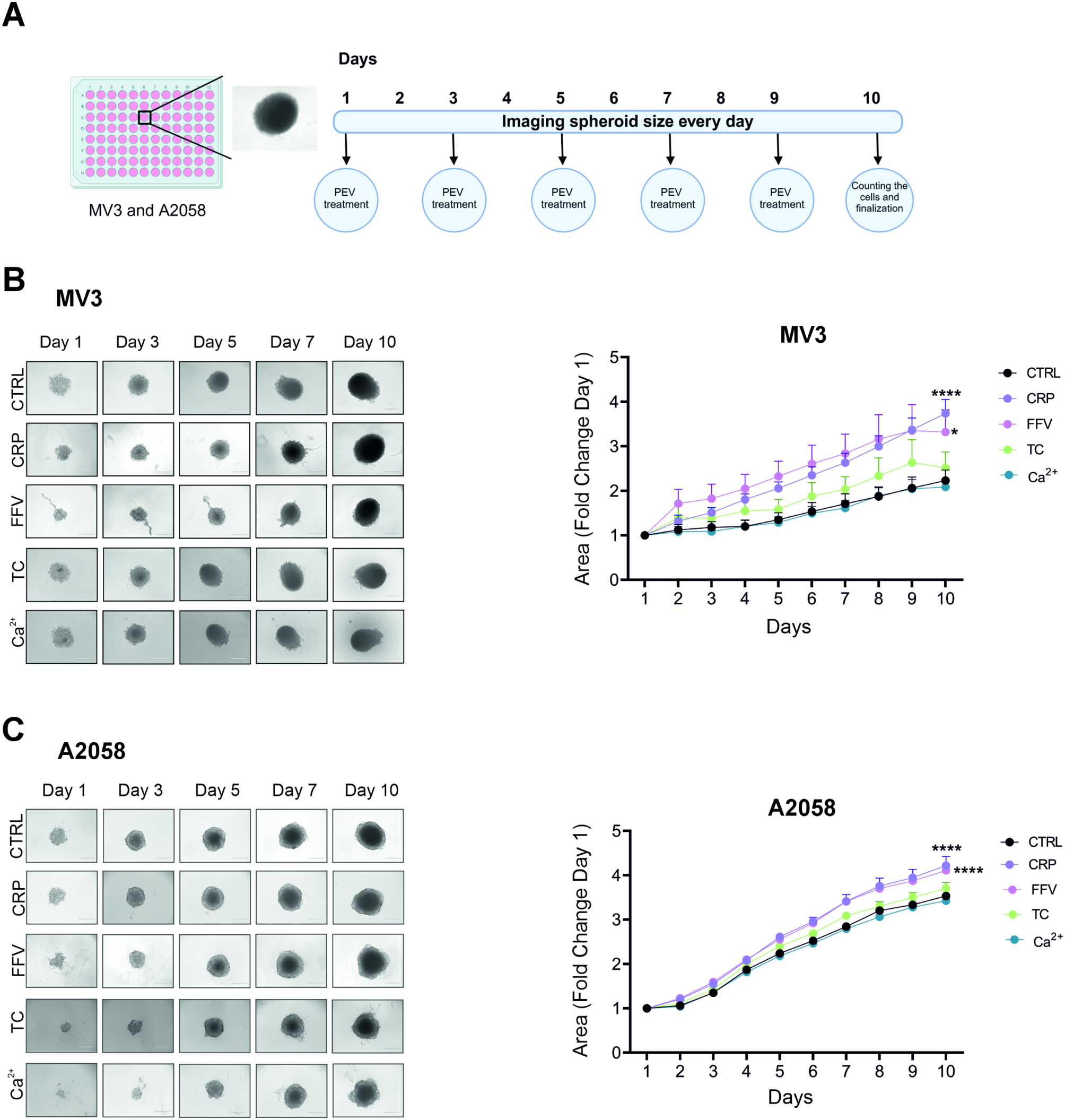
In our hands, undifferentiated NSC-34 cells did not reveal spontaneous neurite growth (Fig. 1A), whereas following differentiation, induced by shifting cells from proliferation to neurobasal medium, NSC-34 cells started to develop neurites that progressively stretched and branched (Fig. 1B). Along with the NSC-34 morphological differentiation, we observed the up-regulation of cytoskeletal marker genes, as evidenced by the increased mRNA levels of MAP2 (microtubule-associated protein), GAP-43 (growth-associated protein 43) and the motor neuron differentiation marker ChAT (choline acetyltransferase) at 4–6 days after the switch to neurobasal medium (Fig. 1C). Moreover, neuron-specific miR-9-3p and miR-124-3p were up-regulated during motor neuron development (Fig. 1D), in accordance with previous studies showing that these two miRNAs are abundantly expressed in the mammalian nervous system and are mainly involved in controlling neuron fate and synaptic morphology [36, 37].
Fig. 1
Quantification and characterization of the EVs released during NSC-34 differentiation. Light microscopy images of the NSC-34 cells before (A; size bar: 200 μm) and after (B; size bar: 100 μm) inducing neuron process outgrowth. In particular, picture B clearly shows neurite development and branching obtained after 4 days of differentiation. Cell viability assayed after 4 days of differentiation is shown in Additional file 10. C) Time course of gene expression analysis of the key differentiation markers MAP2 (microtubule-associated protein), GAP-43 (growth associated protein 43) and ChAT (choline acetyltransferase), and (D) MotomiRs miR-9-3p and miR-124-3p in differentiating NSC-34 cells (n = 3, * p < 0.05, ** p < 0,01; one-way ANOVA test followed by Dunnett’s post-hoc test). The process of differentiation was carried out on NSC-34 cells for two days (Diff-T2), and four days (Diff-T4), and at each of these time points, the collection and characterisation of EVs was undertaken. E) Schematic representation of the serial ultracentrifugation protocol used to harvest EVs from conditioned medium, and separate large from small EVs (lEVs and sEVs, respectively); the NTA distribution plots of vesicle hydrodynamic diameter revealed the difference in size between lEVs and sEVs. F) The size distribution of lEVs and sEVs was also confirmed using transmission electron microscopy (in the panels b, c, d, e and f the size bar corresponds to 100 nm, while in the panels a and g the bar corresponds to 200 nm). G) Western blots analyses show the presence of the well-established EV markers: Alix, Hsp70 and Tsg101, and the negative marker for sEVs, Calnexin (5 µg of total proteins were loaded per lane). Molecular weight markers (kDa) are indicated. Original, uncropped immunoblots are reported in Additional file 7- Fig. S1
Based on the above results, NSC-34 cells were used as a model to study the release of EVs during motor neuron development. To this end, conditioned medium was harvested at various time points of NSC-34 differentiation and EVs were purified using a serial ultracentrifugation protocol precisely adjusted to remove cellular debris and isolate large and small EVs (lEVs and sEVs; Fig. 1E). The nanoparticle tracking assay of purified EVs showed that sEVs had a hydrodynamic diameter of about 122 nm. In comparison, lEVs showed a much more variable size with an average diameter of 170 nm (Fig. 1E). Furthermore, when examined by transmission electron microscopy (TEM) using negative staining, the isolated EVs appeared as closed rounding vesicles delimited by membrane structures; sEVs appeared as vesicles with an outer dense wall and an inner less dense region of approximately 90–120 nm in diameter, whereas lEVs presented a larger and variable size than sEVs of about 140–200 nm in diameter and some of them displayed an electron-dense material delimited by well-defined membrane structure (Fig. 1F). These data are in agreement with common EV features reported in literature [38]. Finally, western blot analysis showed that sEVs are negative for calnexin, a marker of the endoplasmic reticulum and positive for Alix; moreover, sEVs and lEVs are both positive for Hsp70 and Tsg101, two well-defined EV markers (Fig. 1G).
Analysis of EV-miRNAs released during NSC-34 differentiationSince EVs can mediate intercellular communication through their cargo of miRNAs, we performed a small-RNA sequencing experiment to examine the whole miRNA cargo of EVs released during NSC-34 motor neuron development. We analysed three replicates of four EV samples (n = 12), including lEVs and sEVs released at early (Diff-T2) and late (Diff-T5) NSC-34 differentiation time (Additional file 1). A total of 245 miRNAs were detectable in at least two samples. Firstly, the miRNA segregation into the two EV populations, without accounting for the differentiation time, was analysed: of the 245 miRNAs detected, 26 miRNAs (17.9%) were retained only in sEVs, 19 miRNAs (13.1%) were packaged only in lEVs, and 100 miRNAs (69%) were present in both vesicle populations (Fig. 2A, Additional file 4). These data suggest that a fraction of miRNA is specific for either sEVs or lEVs.
Fig. 2
NGS analysis of miRNAs loaded into EVs and released during the NSC-34 differentiation process. A) Descriptive analyses of miRNAs detected in small and large EV samples (N = 12) at two differentiation time points (Diff-T2 and -T5) using small RNA seq. In the Venn diagrams, each area reports the number (%) of miRNAs detected in the corresponding group in at least two samples (N = 3 per group). The upper part of the panel A shows the comparison of the miRNAs detected in lEVs and sEVs regardless the differentiation time, while in the lower part lEVs and sEVs have been analysed separately, and the differences between the number of miRNAs detected at Diff-T2 and Diff-T5 have been reported. B) Unsupervised PCA analysis revealing the similarity between groups. The progressive separation of EV samples collected at Diff-T5 can be appreciated. C) and D) Volcano plot evidencing the number of miRNAs differentially abundant (Moderated t-test < 0.05, fold change > 1.5) in small and large EVs, respectively, when comparing Diff-T5 vs. Diff-T2. E) and F) Heatmap representation of the differentially abundant miRNAs in three EV replicate samples during neuronal differentiation. Red color indicates greater abundance, green color indicates lower abundance. The Panel G reports the list of miRNAs differentially abundant in large and small EVs at Diff-T5 differentiation and their relative expression. The X-axis represents the Log2 fold change in miRNA counts between Diff-T5-lEVs vs. Diff-T5-sEVs. Green bars are miRNAs more abundant in large EVs at Diff-T5, red bars are miRNAs more abundant in sEVs at Diff-T5
Regarding the motor neuron development process, the analysis of the miRNAs released in small and large EVs showed that 60.3% (76 miRNAs) are commonly released at both Diff-T2 and Diff-T5 differentiation times, whereas the remaining miRNAs are differentially abundant in early and late NSC-34 differentiation EVs (Fig. 2A, Additional file 5 and 6).
The unsupervised Principal Component Analysis (PCA) conducted on all samples demonstrated a progressive distance between samples at Diff-T5 and confirmed the differences between the cargo of small and large vesicles (Fig. 2B). We then focused on the identification of the miRNAs differentially abundant in small and large EVs as a function of the differentiation time. We compared Diff-T5 sEVs vs. Diff-T2 sEVs and Diff-T5 lEVs vs. Diff-T2 lEVs. Of the total 126 miRNAs found in the sEVs, 14 miRNAs were significantly more abundant in Diff-T5 and 8 miRNAs in Diff-T2, as evidenced by volcano plot and heatmap analysis (Fig. 2C and E). Instead, of the total 119 miRNAs found in lEVs 30 out of 32 differentially abundant miRNAs were upregulated at the late time of differentiation (Fig. 2D and F). This evidence suggests that the number of secreted miRNAs increases during NSC-34 differentiation, with the most noticeable increase occurring in lEVs compared to sEVs (Fig. 2). This observation was confirmed when we compared the miRNAs significantly modulated in Diff-T5 lEVs vs. Diff-T5 sEVs (Fig. 2G), confirming that miRNA secretion during neuronal differentiation is both stage and EV-type specific.
Functional analysis and gene expression validation of the differentially modulated EV-miRNAsIt has been suggested that miRNAs packaged into EVs can modulate the phenotype of recipient cells. To obtain a picture of the pathways possibly regulated by EV-miRNAs released during NSC-34 differentiation [39], two different web-based analytic tools were used, specifically DIANA miRPath [40] and miRSystem [41]. Moreover, since NSC-34 cells express motor neuron-like features mainly at late differentiation step, only the miRNAs significantly up-regulated in Diff-T5-sEVs and -lEVs were used as a query for the in silico search.
Based on these algorithms, we were able to identify the putative targets of the EV-miRNAs and their enriched pathways. Among the significantly enriched pathways, there are pathways known to have a key role in motor neuron development and intercellular communication, such as axon guidance, neurotrophin, Wnt and TGF-? (Fig. 3A) [42]. To further support these findings, the expression of miRNAs regulating these pathways (miR-9-3p, -16-5p, -335-5p, -669a-3p, -344-3p, -218-1-3p, -124-3p, -709, -34c-3p, -185-5p) were validated by qRT-PCR both in cell bodies and EVs released during NSC-34 differentiation. Quantitative RT-PCR confirmed the NGS results, and most of the analysed miRNAs were up-regulated in NSC-34 cell bodies and EVs along with neurite stretching (Fig. 3B). These data confirm that a number of miRNAs regulated during NSC-34 differentiation and involved in motor neuron maturation are also loaded into EVs suggesting that they could have a role in the crosstalk between motor neurons and surrounding cells.
Fig. 3
Dissecting the complex signals carried by neuron-derived EVs during NSC-34 differentiation. A) miRNAs significantly modulated in lEVs and sEVs collected from late stage of the NSC-34 neuron process outgrowth compared to early stage development using DIANA-mirPath v.3 (https://dianalab.e-ce.uth.gr/html/mirpathv3/). DIANA-mirPath is a miRNA pathway analysis web-server, providing accurate statistics, to predicted miRNA targets (in CDS or 3’-UTR regions) provided by the DIANA-microT-CDS algorithm or even experimentally validated miRNA interactions derived from DIANA-TarBase. In the three panels top ten pathway rankings were reported. B) Since, the prediction analysis of the significantly modulated EV-miRNAs, obtained by NGS, suggested that Axon guidance, neurotrophin, Wnt and TGF-? signalling pathways could be the targets of the miRNAs loaded into NSC-34-derived EVs, qRT-PCR analyses of selected key miRNAs (miR-9-3p, -16-5p, -335-5p, -669a-3p, -344b-3p, -218-1-3p, -124-3p, -709, -34c-3p, -185-5p) were performed both in cell bodies and EVs with the aim to confirm miRNA expression. MiRNA expression analysis performed in lEVs and sEVs at early (Diff-T2) and late (Diff-T5) is reported on the left part of the panel B (n = 3, * p < 0.05, ** p < 0,01; Diff-T5 vs Diff-T2 were compared by t-test for lEVs and sEVs separately); in the right part of the panel B, miRNA expression levels were quantified in cell bodies (n = 3, * p < 0.05, ** p < 0.01; t-test)
NSC-34-derived EVs carry WntsWnts act as morphogens to regulate crucial events of cell fate and development; in the formation of neuronal circuits, Wnts regulate axon guidance, dendritic development, synaptogenesis and synaptic function [43]. More recently, exosomes have emerged as potential carriers for Wnt secretion [14,15,16,17, 44]. Interestingly, NGS analysis of the NSC-34 EV-miRNAs highlighted the loading into EVs of many miRNAs involved in the modulation of axon guidance and Wnt signalling pathway (Fig. 3A and B). Since several studies pointed out the role of the Wnt-associated signalling network in NMJ formation, further experiments were performed to investigate whether NSC-34-derived EVs are loaded with Wnts beyond the miRNAs involved in Wnt signalling modulation.
At first, the mRNA expression levels of several Wnts during NSC-34 differentiation was examined (Fig. 4A). Of note, Wnt1, Wnt3, and Wnt11 were expressed at levels significantly higher than the other tested Wnts, followed by Wnt5a and Wnt9a; on the contrary, the expression of other isoforms, such as Wnt3a, Wnt4, Wnt7b, and Wnt10, was barely detectable. Interestingly, among the most abundant transcripts Wnt1 and Wnt3 increase their expression during NSC-34 differentiation; Wnt5a is the only isoform down-regulated, while Wnt11 is constitutively expressed at high levels. Subsequently, western blotting analyses were performed to investigate whether EVs carry Wnt morphogens into the extracellular environment. The activity of Wnts, Agrin and its coreceptors Lrp4 and MuSK is critical for AChR clustering and thus for NMJ formation. Since in our experimental setup, Agrin protein was barely detectable in NSC-34 cells (Fig. 4B), and it was not found at all in EVs (data not shown), we focused on Wnt4 and Wnt11 which can stimulate clustering, independently from Agrin, directly binding MuSK. Wnt4 is barely detectable both at mRNA level (Fig. 4A) in the whole cell and at protein level in EVs (Fig. 4C; the Wnt4 protein was undetectable in the whole cell, data not shown) [45]. Whereas the other Agrin-independent stimulating Wnt, Wnt11 protein is easily detectable in NSC-34 cell bodies as shown in Fig. 4B, and in both lEVs and sEVs (Fig. 4C). Finally, we evaluated Wnt3a protein expression, one of the best characterised Wnts. As shown in Fig. 4B, Wnt3a shows a decreasing trend during neurite development, and in our hand, it is loaded into sEVs but not lEVs mirroring a reduction during NSC-34 differentiation (Fig. 4C). To confirm Wnt loading into EVs further, well-established markers (Tsg101, CD9 and CD81 and the negative control for sEVs, Calnexin; Fig. 4C) were used to characterise lEVs and sEVs. The obtained results suggest that EVs could modulate NMJ development through Wnts.
Fig. 4
Expression analysis of Wnts during NSC-34 differentiation. (A) Gene expression analysis of Wnt isoforms in NSC-34 whole cell during neuron process outgrowth (Diff-T0: undifferentiated NSC-34 cells; Diff-T2: NSC-34 switched to neurobasal medium for 2 days; Diff-T5: NSC-34 switched to neurobasal medium for 5 days; n = 3, * p < 0.05, ** p < 0,01; one-way ANOVA test followed by Dunnett’s post-hoc test). (B) Western blot analysis of Agrin, Wnt3a, and Wnt11 in differentiating NSC-34 cells. In each lane 30 µg of total proteins were loaded and actin expression was used as loading control. (C) lEVs and sEVs were isolated from NSC-34 during differentiation as indicated in Fig. 1. Whole cell extracts of undifferentiated NSC-34 cells (CB Diff-T0) and extracellular vesicles (lEVs and sEVs) at indicated times were probed with Tsg101, CD81 and CD9, used as EV markers, and Calnexin as the negative control. The figure clearly shows that Wnt11 is present in both lEVs and sEVs whereas Wnt3a is expressed only in sEVs, with a decreasing trend during neurite process development. Five µg of total proteins were loaded in each lane. Molecular weight markers (kDa) are indicated. Original, uncropped immunoblots are reported in Additional File 8 - Fig. S2
NSC-34-derived EVs stimulate C2C12 plasma membrane remodelling favouring AChR clusteringAn accurate communication process between the motor neuron and the skeletal muscle fibre is required for proper NMJ assembly, growth, and maintenance. Based on the above-reported observations that either miRNAs or Wnt11, both involved in NMJ formation, are loaded into NSC-34-derived EVs, we investigated whether the EVs from NSC-34 cells (donors) can stimulate plasma membrane remodelling in C2C12 myotubes (targets). To address this question, a transwell co-culture system with C2C12 myotubes (bottom) and differentiated NSC-34 (top) was set up (Fig. 5A); after two days of co-culture, the signals released into the environment, including EVs, induced R-Spondin 2 (Rspo2) upregulation and Acetylcholine receptor alpha (AChR alpha) downregulation in myotubes, while ß-catenin and Vangl2 were unchanged. These data suggest that NSC-34 and C2C12 cross-talk can lead to myotube membrane remodelling, an initial phase of NMJ formation (Fig. 5B).
Fig. 5
Motor neuron-derived EVs stimulate AChR cluster formation in C2C12 plasma membrane. (A) C2C12 myotubes and differentiating NSC-34 cells were co-cultured and allowed to interact for 48 h without physical contact thanks to the presence of a membrane filter of 0.4 μm pores interposed between the two cell lines. (B) After 48 h co-culture, myotubes were harvested and mRNA extracted to evaluate the gene expression of selected targets; the obtained data show that the presence of NSC-34 cells (and likely their released signals) induced in C2C12 myotubes a decrease and an increase in AChR alpha and Rspo2 mRNAs, respectively (n = 3, * p < 0.05; t-test). (C) The α-Bungarotoxin confocal laser scanning microscopy analysis highlights a significant increase in AChR clusters in response to the EV administration. (D) The morphometric analysis of the AChR clusters shows that lEV and sEV treatments increased the number of AChR clusters per µm of fibre and reduces the cluster area; in detail the increase of the fluorescence intensity emitted from clusters in the treated samples, is more evident when the intensity was related to the cluster area (* p < 0.05, one-way ANOVA test followed by Dunnett’s post-hoc test). (E) At the molecular level, lEVs and sEVs induce the stimulation of Rspo2 in the EV-treated myotubes and a tendency to decrease of AChR alpha and Axin-1 mRNA expression (* p < 0.05, one-way ANOVA test followed by Dunnett’s post-hoc test). (F) An ex vivo model of primary motor neurons derived from spinal cord explants was used to corroborate the results obtained using the NSC-34 model. Total EVs were collected from motor neurons grown for 9–11 days after explantation and used to treat mature myotubes; the conditioned medium (CM) not centrifuged was used to compare bulk with vesicular signals. (G) In line with NSC34-derived EV effects, Rspo2 and Axin-1 mRNA levels are modulated by motor neuron EVs (* p < 0.05, one-way ANOVA test followed by Dunnett’s post-hoc test for Rspo2 and Bonferroni post-hoc test (Medium vs. EVs) for Axin-1)
Subsequently, to gain insights into the role of NSC-34-derived EVs on the modulation of muscle membrane receptors, an event that precedes the juxtaposition of motor neuron termination to the muscle fibre, C2C12 myotubes were treated with EVs purified from five-day differentiated NSC-34 cells (Diff-T5 lEVs and sEVs). Thus, we aimed to evaluate whether NSC-34-derived EVs could modulate the levels of AChR clustering in myotubes.
Immunofluorescence staining with α-Bungarotoxin (Fig. 5C) allowed us to highlight a significant increase in AChR clusters in response to the EV administration. Morphometric analysis of the AChR clusters showed that lEV and sEV treatments increased the number of AChR clusters per µm of fibre and reduced the cluster area (Fig. 5D). The reduction of cluster area can be explained as a lower diffusion of AChRs on the myotube plasma membrane due to clustering. Finally, the fluorescence intensity emitted from clusters was higher in the treated samples, more clearly when the intensity was related to the cluster area (Fig. 5D). These data are in agreement with the stimulation of Rspo2 in the EV-treated myotubes, whereas AChR alpha and Axin-1 mRNA expression showed a tendency to decrease (Fig. 5E). Axin-1 and Rspo2 modulate Wnt signalling supporting the hypothesis of a possible involvement of the Wnt pathway in stimulating AChR clustering.
According to the results obtained using the NSC-34 in vitro model, we attempted to corroborate our findings using an ex vivo model of primary motor neurons derived from spinal cord explants (Fig. 5F). The EVs were collected from motor neurons grown for 9–11 days after explantation. In detail, conditioned media were collected and ultracentrifuged following a similar protocol used to isolate NSC-34-derived EVs. Still, we no longer separated EVs in different sub-populations due to the low amount of collected EVs and since NSC-34-derived lEVs and sEVs induced overlapping effects on myotubes.
Then, mature myotubes were treated both with the EVs isolated from the ex vivo model of primary motor neurons and motor neuron conditioned medium not centrifuged (CM, soluble and vesicular signals) (Fig. 5F). As shown in Fig. 5G, in line with NSC-34-derived EVs effects, Rspo2 mRNA levels were higher in C2C12 treated with conditioned medium and EVs than in control samples, while Axin-1 expression seems to be slightly down-regulated in response to EV treatment; conversely, it increased following the treatment with not centrifuged medium. Moreover, Rapsyn expression was not modulated in response to both EV treatment and to not centrifuged medium (Fig. 5G).
Altogether, the collected data demonstrate that motoneurons release EVs containing molecular signals, proteins and microRNAs able to stimulate AChR aggregation on the myotube membrane, a preparatory stage of the NMJ formation.
NSC-34-derived EVs act as platforms of integrated molecular signals triggering β-catenin dependent and independent Wnt pathways for AChR clusteringThe above reported NGS data showed that EVs secreted from NSC-34 cells are enriched in miRNAs synergically targeting the effectors of the β-catenin dependent Wnt pathway (such as Axin-1, GSK3β, CSNK2a2/1a1, APC and, Ppp2R5c) and leading to its activation (Fig. 6A). Furthermore, we demonstrated that NSC-34-derived sEVs shuttle high levels of Wnt11, a Wnt isoform known to stimulate both the canonical and non-canonical Wnt pathways, and low quantities of Wnt3a, a negative modulator of the Wnt signalling [10, 18, 19]. This evidence prompted us to investigate whether sEV administration to C2C12 cells could activate Wnt signalling contributing to AChR clustering.
Fig. 6
NSC-34-derived EVs trigger β-catenin dependent and independent Wnt pathways for AChR clustering. (A) Target prediction of NSC-34-derived EV miRNAs and DIANAmirPath analysis of Wnt signalling (Kegg pathway), based on DIANA miRPath v.3.0 program. The miRNAs selected from the first two quartiles of Diff-T 2-T5 lEVs and sEVs have been used for the analysis. The miRNAs highlighted in red target the genes involved in the destruction complex of the ?-catenin. (B) The α-Bungarotoxin confocal laser scanning microscopy analysis highlights a significant increase in AChR clusters in response to the Agrin and sEV treatments. (C) Expression analysis of some targets of the Wnt signalling pathway by qPCR analyses in C2C12 myotubes treated with Agrin or increasing quantities of NSC-34 sEVs (sEV concentrations were 5 × 108, 1 × 109, and 5 × 109 particles/mL indicated by symbols). * p < 0.05, one-way ANOVA test followed by Dunnett’s post-hoc test. (D) Western blotting analysis: the increased expression of GSK3 inactive form and JNK active form was measured as ratio of the phosphorylated isoform/not phosphorylated isoform. The fold change refers to the control condition (non-treated C2C12 myotubes). ** p < 0.01, *** p < 0.001, one-way ANOVA test followed by Dunnett’s post-hoc test. (E) Immunofluorescence staining of C2C12 myotubes treated with NSC-34-derived sEVs, representative pictures showing the increased nuclear localization of ?-catenin (red) and DAPI (blue) following EV treatment. (F) Western blotting analysis of the ?-catenin active form in C2C12 myotubes treated with NSC-34 sEVs ranging from 108 to 5 × 1010 particles/mL. Tubulin expression was used as loading control. Arbitrary units refer to the control condition. * p < 0.05; ** p < 0.01 *** p < 0.001, one-way ANOVA test followed by Dunnett’s post-hoc test. Original, uncropped immunoblots are reported in Additional File 9 - Fig. S3
Since Wnt11 can stimulate AChR clustering even in the absence of Agrin, we first administered Agrin or Diff-T5 NSC-34-derived sEVs to C2C12 myotubes, then AChR clustering was studied. Immunofluorescence staining with α-Bungarotoxin showed AChR cluster formation in the presence of both Agrin and sEVs (Fig. 6B), suggesting that Wnt11 carried by sEVs could directly activate the MuSK receptor and stimulate down-stream signalling leading to AChR clustering.
Moreover, the gene expression analysis by qRT-PCR (Fig. 6C) highlighted Axin-1 and β-catenin mRNA down-regulation. Axin-1 is a component of the destruction complex of β-catenin together with adenomatous polyposis coli (APC), glycogen synthase kinase 3 beta (GSK3β) and casein kinase (CSNK), indeed Axin-1 mRNA degradation is compatible with β-catenin-dependent Wnt activation and miR-124-3p activity. Of note, Rspo2 increased after sEV treatment further corroborating Wnt-dependent AChR clustering (Fig. 6C) [46]. The immunofluorescence analysis evidenced a clear translocation of β-catenin into the nucleus of myotubes treated with NSC-34-derived sEVs (Fig. 6D) Moreover, western blotting analysis of β-catenin active form further supported the sEV positive stimulation of the canonical Wnt signalling (Fig. 6E). This evidence is in agreement with the accumulation of p-GSK3β following treatments with increasing concentration of the NSC-34-derived sEVs (Fig. 6F).
Since Wnt11 could also activate non-canonical Wnt/PCP pathway through the hetero-dimer Lrp4/MuSK, the mRNA expression of Lrp4 and Lrp5/6 was also investigated; qRT-PCR data showed Lrp6 down-regulation, while Lrp4 mRNA was unchanged suggesting that the formation of the hetero-dimer receptor Lrp4/MuSK could be favoured in response to EV treatment (Fig. 6C). Western blot analysis of JNK, one of the down-stream effectors of MuSK, revealed an increase in p-JNK in response to NSC-34-derived sEVs (Fig. 6F).
The results obtained support the current literature describing the involvement of both canonical and non-canonical signalling pathways in NMJ formation; in fact, the data reported here show that EV treatment induces AChR clustering and this phenomenon is associated with β-catenin translocation into the nucleus, suggesting activation of canonical Wnt signalling. Furthermore, as suggested by the increase in p-JNK following EV treatment, it appears that NSC-34-derived EVs could also activate non-canonical signalling pathways thanks to their complex cargo. Interestingly, EV miRNAs appear to down-regulate Lrp6 mRNA, consequently Lrp6 mRNA decrease could favour the formation of the Lrp4/Musk receptor dimer, further supporting the idea of the activation of Wnt11-Lrp4/Musk-JNK axis.
EVs carrying Wnt11 induce GSK3β phosphorylationPrevious experiments suggested that motor neuron-derived EVs may contribute to the early stages of NMJ development by inducing the clustering of AChRs through the activation of canonical and non-canonical Wnt signalling pathways. In this scenario, we hypothesised that Wnt11 could act as an upstream stimulus, while the miRNAs enriched in NSC-34 EVs could fine-tune the downstream effectors of Wnt11. In an attempt to dissect the role of Wnt11 and miRNAs in this process, we forced the expression of exogenous Wnt11-HA tag fusion protein in Hek293 cells, then we collected sEVs from transformed Hek293 and used them to treat myotubes. After 18 h treatment, the modulation of Wnt signalling was investigated. First, we performed NTA of sEVs isolated by serial ultracentrifugation (Fig. 7A), then using western blot analysis we showed that the Hek293 cells overexpressing Wnt11-HA tag fusion protein secreted sEVs positive for CD63, a typical exosomal marker, and Wnt11 (Fig. 7B). Although, sEVs from wild-type Hek293 show the expression of low levels of endogenous Wnt11, the positivity for HA tag only in transformed Hek293 cells unambiguously demonstrates the loading of exogenous Wnt11-HA tag into sEVs (Fig. 7B). Notably, this evidence extends the finding that EVs carry Wnt11. Subsequently, EVs loaded with exogenous Wnt11 but not motor neuron-derived miRNAs were administered to myotubes, and the WNT pathway was studied. Figures 7C and D clearly show that the administration of Hek-Wnt11 sEVs to myotubes induced GSK3β phosphorylation; interestingly a slight increase in GSK3β phosphorylation was also observed for wt Hek sEVs (Fig. 7C and D), probably due to the presence of low levels of endogenous Wnt11. Evaluation of downstream markers of canonical Wnt signalling revealed that, although Wnt11 led to the GSK3β phosphorylation, the corresponding activation of β-catenin was not detected (Fig. 7C). These findings suggest that, in our experimental setup, NSC-34-derived miRNAs seem to be crucial for an efficient β-catenin activation. Moreover, Hek-Wnt11 sEV administration to myotubes did not induce a significant JNK phosphorylation (Fig. 7C and D). Overall, these data suggest that, without NSC-34 miRNAs, Wnt11 has a negligible effect on activating the non-canonical Wnt signalling pathway. However, it appears to activate GSK3β phosphorylation, a typical step in the canonical pathway.
Fig. 7
Hek-WNT11 sEVs induce GSK3β phosphorylation. (A) NTA distribution plot of the hydrodynamic diameter size of Hek-WNT11 sEVs. (B) The Western blot analysis of the sEVs secreted by wild type and WNT11-HA tag-overexpressing HEK cells shows the presence of the well-established exosomal marker CD63, and the WNT11 or HA tag (10 µg of total proteins were loaded per lane). Molecular weight markers (kDa) are indicated. (C) Western blot analysis of GSK3α/β, JNK and β-catenin in C2C12 myotubes treated with increasing concentrations of Hek sEVs (lanes 2–4) or Hek-WNT11 sEVs (lanes 5–7) as indicated in the figure. Non-treated C2C12 myotubes represent the control (lane 1); Tubulin was used as the housekeeping protein. Molecular weight markers (kDa) are indicated. Original, uncropped immunoblots are reported in Additional File 11 - Fig. S4. (D) Expression of GSK3 inactive form e JNK active form was measured as ratio of the phosphorylated isoform/not phosphorylated isoform. The fold change refers to the control condition. *** p < 0.001, one-way ANOVA test followed by Dunnett’s post-hoc test
Comments (0)