
Remember me


The diet-induced obesity model (DIO) was carried out with female BALB/c mice, which were fed on an HFD for 12 weeks, as previously described (Fig. 1A) [16]. The diet consisted of approximately 21% metabolizable energy (ME) from carbohydrates, 19% from protein, and 60% from fat, signifying higher fat and lower carbohydrate content compared to the NFD (62% carbohydrates, 20% protein, 5% fat) (Supplemental Tab. S1, Supplemental Fig. S1A). Throughout the dietary period, body weight was monitored weekly, and fecal pellets were collected before and after the diet (Fig. 1A). The relative body weight showed a significant increase in the cohort of mice on the HFD (Fig. 1B). Moreover, mice on the HFD exhibited a notable increase in intra-abdominal adipose tissue mass and blood glucose levels (Fig. 1C).
Fig. 1
HFD results in a distinct obese phenotype with a shift in gut microbiome composition. Schematic representation of the in vivo diet-induced obesity (DIO) model with a 12-week HFD and collection of fecal pellets before and after the dietary period (A). The 12-week dietary period increased relative body weight (B) (NFD: n = 21, HFD: n = 20) with higher intra-abdominal adipose tissue mass (NFD: n = 6, HFD: n = 6) and blood glucose levels after the diet (NFD: n = 21, HFD: n = 20) (C). Values in panels B and C are displayed as mean ± SD. Gene correlation after shotgun metagenomics sequencing of fecal pellets shows correlation coefficients pre-diet and post-diet (D). HFD caused a taxonomical shift in gut microbiome composition at grouped phylum (E) and genus (F) levels, displayed as heatmaps with each group (n = 3). Additionally, it altered the functional profile after genetic annotation to the KEGG database level 1 (G) and CaZy level 1 (H), shown as absolute value of “Z” representing distance from raw value to population mean in SD units. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. P calculated by 2-way ANOVA Šídák’s multiple comparisons test (C) and Mann-Whitney test (D)
To study the effects of the HFD on the microbial environment of the gut, we conducted shotgun metagenomic sequencing using fecal samples, resulting in information about the taxonomic variety of microbes and their functions. We analyzed fecal samples before starting the diet (pre-diet) and after 12 weeks of either NFD or HFD (Fig. 1D) and found that all mice receiving the same diet exhibited a positive correlation in gene expression. Notably, mice that transitioned from NFD to HFD displayed an even stronger positive correlation, with a mean correlation coefficient of 0.585, compared to NFD mice, which had a mean correlation coefficient of 0.365 (Fig. 1D).
The analysis of genes in the mice cohort showed that those receiving the HFD had more bacteria from the phyla Firmicutes and Proteobacteria and fewer from Bacteroidetes (Fig. 1E). Additionally, the numbers of bacteria from the Bacteroides, Prevotella, and Lactobacillus genera decreased, while Anaerotruncus increased (Fig. 1F). Further analysis of the murine fecal pellets revealed differences in gene abundance between the HFD and NFD cohorts. Genes related to general metabolism, organismal systems, genetic information processing, and human diseases were less abundant in the HFD mice, while genes linked to environmental information processing and cellular processes were more abundant (Fig. 1G). Comparing the abundant genes with the CAZy database showed that the HFD group had reduced expression of genes related to carbohydrate-active enzymes compared to the NFD group (Fig. 1H).
Hence, feeding mice with HFD not only resulted in an increase in body weight, abdominal adipose tissue mass, and blood glucose levels. Moreover, this specific diet led to taxonomical and functional shifts in the microbial community within the gut.
Elevated virus titers and increased pulmonary inflammation in HFD mice infected with IAVAfter the dietary period, mice were infected with an H1N1 IAV strain, obtained from an obese patient at the Jena University Hospital. Before infecting NFD and HFD mice, the virus strain was modified through passaging, plaque purification, and isolation, as previously described [16]. Mice from both dietary groups (w or w/o IAV) were euthanized 2 dpi to assess the acute phase of infection, and 21 dpi to evaluate long-term changes in mice after (Fig. 2A). Throughout the infection period, all mice were monitored daily, and body weight and burden score values were recorded. The scoring system not only considers body weight but also includes additional parameters to assess the severity of the disease in mice infected with IAV [16]. The burden score values for NFD mice decreased from day 5, reaching an average maximum at day 4 p.i. Interestingly, HFD mice reached maximal score values at day 4 p.i. and maintained them at the same level until day 10 p.i. (Fig. 2B). H&E staining was performed on the left lung lobe of mice both before infection and at 2 and 21 dpi (Supplemental Fig. S1B). In addition to the qualitative observations of pulmonary inflammation within the tissue, a histoscoring system was utilized to quantify these findings (Supplemental Tab. S2). At both time points, the infection led to an increased histoscore; however, no clear trend was observed between mice on NFD and on a HFD (Supplemental Fig. S1C). At 2 dpi, the number of infectious virus particles detected in homogenized lung tissues was significantly higher in mice on a HFD (Fig. 2C). Additionally, the levels of pro-inflammatory mediators in the lung homogenates of infected mice at 2 dpi were elevated in HFD mice, particularly interferon (IFN)-α, IFN-γ, and interleukin (IL)−1β (Fig. 2D).
By day 21 p.i., no infectious virus particles were found in the lungs of either the non-obese or obese mice. However, lung tissue from HFD mice produced significantly higher amounts of IP-10 (encoded by CXCL10) and IL-1β. The levels of the interferons, IFN-α and IFN-γ, remained elevated in the lungs of HFD mice three weeks after infection (see Fig. 2E). Moreover, the levels of IL-10, C-X-C Motif Chemokine Ligand 1 (CXCL1), Regulated and Normal T cell Expressed and Secreted (RANTES), and Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) were predominantly increased in the infected HFD group (Supplemental Fig. 2). Interestingly, a similar trend toward increased pulmonary inflammation was also noted in uninfected HFD mice, although it did not reach statistical significance (Fig. 2E).
In summary, infection of HFD mice led to significantly increased virus loads in the pulmonary system accompanied by increased inflammation at an acute state of infection and after three weeks.
Fig. 2
Elevated virus titers and increased pulmonary inflammation in HFD mice infected with IAV. Schematic overview of intranasal infection of NFD (n = 15) and HFD (n = 14) mice with IAV, including organ harvest and fecal pellet collection after 2 and 21 days (A). Scoring values for NFD and HFD mice across 3 weeks p.i. (B). HFD mice (n = 6) exhibited higher active virus particles in lung tissue at day 2 p.i. compared to NFD mice (n = 3) (C). HFD mice showed altered pulmonary cytokine profiles at acute phase (2 days post infection (dpi), HFD n = 6, NFD n = 3) (D) and long-term (21 dpi, HFD n = 6, NFD n = 6) (E) compared to mock NFD (n = 6) and HFD (n = 6) mice. Values are mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, calculated by Kruskal-Wallis test, Dunn’s multiple comparison test (C-E)
Dysregulated gut microbiome taxonomy and SCFA production in infected HFD miceThe correlation analysis of bacterial genes isolated from the fecal pellets of both dietary cohorts of uninfected and infected mice at day 2 and 21 p.i. showed a similar pattern to the one observed after an HFD (Fig. 3A). Furthermore, the increased ratio of Firmicutes to Bacteroidetes, previously observed in mice after the HFD, was still present during infection. Additionally, the phylum Proteobacteria was increased in HFD mice before, during acute infection, and after 21 days. Bacteria of the phylum Verrucomicrobia were predominantly increased in infected obese mice after 2 days (Fig. 3B).
Fig. 3
Dysregulated gut microbiome taxonomy and SCFA production in infected HFD mice. Gene correlation analysis after shotgun metagenomics from fecal pellets at 2 and 21 days post infection (dpi) from NFD and HFD depends on diet (A). Changes in microbiota composition during infection depend on diet at the phylum level (B). Heatmaps from LefSe analysis show significantly different genera related to SCFA production between HFD (n = 3) and NFD (n = 3) mice at mock, 2 dpi, and 21 dpi. The heatmap values are standardized Z values for relative abundance (C). Reduced expression of Acetate kinase (EC 2.7.2.1) was significant in mock mice after 12 weeks of diet, shown as relative abundance (NFD, n = 3; HFD, n = 3). *P < 0.05 & *Q < 0.05 calculated via Metastats (D)
We studied the impact of diet and IAV infection on specific metabolite-producing bacteria. For this, we focused on bacterial genera known to produce SCFA such as acetate, propionate, and butyrate before and during infection, as described by Koh et al. [23]. LefSe (linear discriminant analysis (LDA) Effect Size) was used to identify genomic features (genes, pathways, or taxa) that characterize the differences between various conditions (Fig. 3C). In the absence of infection, HFD mice displayed reduced levels of Prevotella, Bacteroides, and Lactobacillus bacteria, which are known to produce acetate and propionate. However, butyrate-producing genera Roseburia and Faecalibacterium increased. Bacteroides and Ruminococcus, which also produce acetate and propionate, decreased in obese mice at day 2 and 21 p.i. Furthermore, Prevotella, which can also produce acetate, decreased. Faecalibacterium bacteria, known for producing butyrate, increased at day 21 p.i. (Fig. 3C).
During the infection, genes related to lipid metabolism remained reduced in mice on a HFD compared to mice fed with a NFD. Interestingly, genes associated with human diseases increased in HFD mice during the acute phase of infection, at 2 dpi (Supplemental Fig. S3A). Analysis of CAZy showed a similar reduction as seen after the diet, with a decreased expression of genes in all six categories (Auxiliary Activities (AA), Carbohydrate Binding Molecules (CBM), Carbohydrate Esterases (CE), Glycoside Hydrolases (GH), Glycosyl Transferases (GT), Polysaccharide Lyase (PL)) in HFD-receiving mice, at 2 dpi and 21 dpi, except for AA, which was comparatively increased at day 2 p.i. (Supplemental Fig. S3B).
Despite the limited volume of serum available in mice for SCFA detection, we successfully quantified acetate levels in a reduced number of uninfected normal-fat diet (NFD) and high-fat diet (HFD) mice after the dietary period. Although the levels were decreased, this change did not reach statistical significance (Supplemental Fig. S3C). To evaluate genes related to the production of SCFAs, we conducted a KEGG analysis focused on SCFA-producing pathways, represented as relative abundances (Supplemental Fig. S3D, Fig. 3D). Notably, the phosphate acetyltransferase-acetate kinase pathway, which is responsible for acetate synthesis, showed a significant reduction in uninfected mice on HFD (Supplemental Fig. S3D). This effect was also observed during infection, although it did not reach statistically significant levels. Additionally, at the enzymatic level, acetate kinase—a key enzyme involved in acetate synthesis—exhibited similar trends (Fig. 3D).
Therefore, after HFD and during infection, the gut microbiome of HFD mice showed alterations at a taxonomical and functional level. Specifically, the production of SCFAs differed, with the relative abundance of genes associated with acetate synthesis being lower in mice on the HFD.
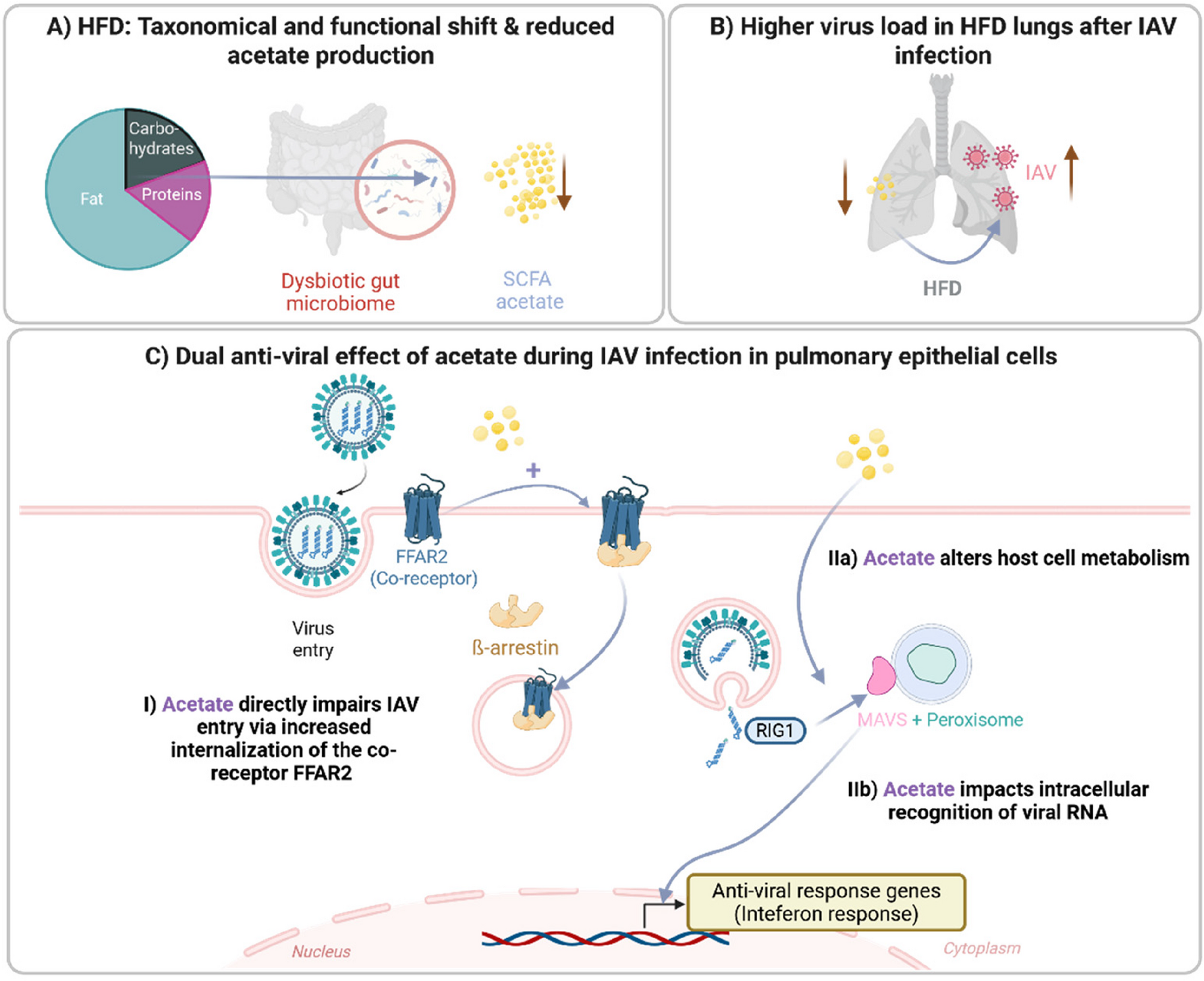
Reduction in virus titer in infected human ex vivo lung slices pre-treated with acetateIn order to analyze the relationship between diet and gut metabolites in relation to infection in humans, we studied the blood of healthy female donors, categorizing them as non-obese or obese based on their BMI [24]. Donors with a BMI of < 25 were classified as “non-obese,” while individuals with a BMI of > 30 were classified as obese [25] (Supplement Tab. S3). Elevated BMI was associated with higher higher serum leptin levels and increased age (Fig. 4A, Supplemental Fig. S4A). Donors were carefully selected within the same age range (30–45 years) to limit age-related variability. Besides leptin, also other cytokines and adipocytokines were determined in the serum, but with no significant trends (Supplemental Fig. S4B). Additionally, we measured levels of the SCFAs acetate, butyrate, and propionate. Acetate exhibited the most notable decrease in the blood of obese donors compared to non-obese donors (Fig. 4B).
Fig. 4
Reduction in virus titer in infected human ex vivo lung slices pre-treated with acetate. Healthy female donor cohorts (according to Supplemental Tab. S3) were characterized by a normal BMI (< 25 (kg/m²), n = 4) and an increased BMI for obese individuals (> 30, n = 6) and elevated systemic leptin levels (A). There was a trend towards reduced systemic acetate (p = 0.2189, ?2 = 0.3376), butyrate (p = 0.6727, ?2 = 0.02703), and propionate (p = 0.4392, ?2 = 0.1009) levels (B). Schematic overview of human ex vivo lung slices (PCLS) treated with SCFAs for 2 days and subsequent infection with IAV for 2 days (C). Acetate treatment of lung slices resulted in lower active virus particles, or plaque forming units (PFU/ml) in supernatants (D). Solely IAV infection increased levels of IFN-α2, IFN-γ, and TNF-α, while acetate pretreatment slightly enhanced cytokine production (E). Reduced IAV matrix protein (green) expression in infected lung slice pre-treated with acetate within the alveolus, co-stained with CD68 (red), DAPI (blue), and Phalloidin (purple) (F). Scale bar = 100 μm. Values in panels A, B, D, and E are mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, P calculated by Mann-Whitney test (A, D), unpaired t-test with Welch’s correction (B), one-way ANOVA, Tukey’s test (E)
To study the potential positive effect of SCFAs on the human lung, we used an already established human ex vivo PCLS model [19]. The lung slices were pre-incubated with specific concentrations of individual SCFAs (acetate, butyrate, and propionate) for 2 days and then infected with an H1N1 strain (PR8/34) for additional 2 days. After the infection, SCFAs were reintroduced into the medium.(Fig. 4C). Interestingly, only the lung slices stimulated with acetate showed a significant reduction in the number of infectious virus particles. The treatment with butyrate and propionate did not show any significant alterations (Fig. 4D). These findings were consistent with an early significant increase in the cytokines IFN-α2, IFN-γ, and tumor necrosis factor (TNF) - α due to infection. Slices additionally treated with acetate did lead to a slight increase of IFN-α and IFN-γ, yet reaching no significant level (Fig. 4E). To confirm these findings, we performed immunofluorescence staining, which also showed a qualitative reduction in virus particles in the alveoli after treatment with acetate in combination with infection (Fig. 4F).
Thus, obese, human, female donors also displayed dysregulated levels of SCFAs in their circulation, with the most noticeable impact seen in reduced acetate levels. This particular SCFA, when analysed during treatment in a human ex vivo system in combination with infection, demonstrated the most significant beneficial antiviral effect.
Antiviral effect of acetate depends on FFAR2 activation and internalization dynamics in pulmonary epithelial cellsTo elucidate the molecular mechanisms underlying the detected antiviral effect of acetate observed during IAV infection in the human lung ex vivo slices, we conducted similar stimulation and infection experiments in pulmonary epithelial cells (A549) for 8 h and 24 h of infection (Fig. 5A). The A549 cell line is known to mimic properties of the alveolar type II cell line, including the presence of lamellar bodies [26]. Stimulation with medium containing 260 µM acetate did not increase cytotoxicity at either 8–24 h (Supplemental Fig. S5A). Consistent with the results from ex vivo lung slices, acetate stimulation reduced virus titer determined in supernatants of A549 cells at both 8 and 24 h post infection (hpi) (Fig. 5B).
Fig. 5
Antiviral effect of acetate depends on FFAR2 activation and internalization dynamics in pulmonary epithelial cells. Schematic representation of A549 cells pre-treated with acetate and infected with IAV for 8 and 24 h (A). Data represent n = 9 independent biological replicates per condition. Acetate treatment reduced active virus particles in the supernatant of infected wildtype A549 at 8 and 24 h post infection (hpi) (B). Acetate treatment increased ß-arrestin2 recruitment to the cell membrane (arrows) in HEK293 cells expressing FFAR2 compared to untreated and FFAR2-deficient cells (C). Furthermore, acetate stimulation caused FFAR2 translocation from the plasma membrane (CAAX) to early endosomes (FYVE) (left), while ß-arrestin2 also translocated to early endosomes (right) (D). Reduction of active virus particles was absent in A549 cellswith global FFAR2 receptor knock-out (E). IP-10 levels were increased in response to infection in supernatants of both A549 WT and FFAR2-KO cells, with a noticeable increase in WT cells at 24 hpi with IAV (n = 3) (F). Acetate pre-treatment on A549 with global knock-out of IP-10 did not affect the number of active virus particles after 8 or 24 hpi (n = 9) (G)
To further analyze the effect of acetate on the FFAR2 receptor, we examined the impact on receptor internalization in HEK293 cells, focusing on the ability to recruit β-arrestin2, an important mediator of GPCR endocytosis [27]. Live-cell confocal microscopy imaging revealed that β-arrestin2 is recruited to the cell membrane in an acetate-dependent manner. This recruitment depends on the presence of the FFAR2 since cells not expressing the receptor do not show any acetate-induced recruitment of β-arrestin2 to the plasma membrane (Fig. 5C).
To comprehensively investigate the internalization and early trafficking characteristics of the FFAR2 we conducted BRET assays utilizing the energy transfer between NanoLuc-tagged FFAR2 or β-arrestin2 and membrane-localized CAAX-YFP (prenylation sequence of KRas, plasma membrane) and FYVE-mNeonGreen (phosphatidylinositol 3-phosphate (PI3P) binding motive of endofin, early endosomes) constructs, as previously described [28]. The analysis revealed an acetate-dependent, rapid translocation of the FFAR2 from the plasma membrane (CAAX) to the early endosomes (FYVE). About ten minutes after acetate treatment, the relocation of FFAR2 to the plasma membrane reaches a plateau that does not return to the basal level, indicating reduced FFAR2 levels at the cell surface in the presence of acetate. Additionally, a part of the internalized FFAR2 stays at the early endosomes 30 min after acetate stimulation. In order for the FFAR2 to be internalized, adaptor protein β-arrestin2 is recruited to the plasma membrane (CAAX) and translocates to the early endosomes (FYVE), most probably together with the FFAR2 (Fig. 5D).
To analyze the effect of acetate and FFAR2 in the pulmonary epithelial cells during IAV infection, we repeated the stimulation and infection setup using A549 cells with a global knock-out of FFAR2 (FFAR2-KO). Interestingly, no reduction of virus titer was observed in acetate-treated FFAR2-KO cells, indicating that the antiviral effect of acetate during IAV infection is dependent on the GPCR FFAR2 (Fig. 5E).
Moreover, we aimed to investigate the possible influence on the interferon system, a crucial player in the antiviral response, by measuring the amount of IP-10 (encoded by CXCL10). This chemokine is known to be produced in response to influenza virus infection and plays a role in modulating the antiviral response [29]. In supernatants of both in vitro cell lines A549 WT and FFAR2-KO, infection with IAV resulted in a significant upregulation of IP-10 after 24 h of infection. Acetate treatment led to slightly increased levels of IP-10, although this increase was not statistically significant (Fig. 5F). In FFAR2-KO cells, this treatment did not show this trend. To assess the impact of the production of IP-10 on the reduced viral titer detected in A549 WT cells, absent in FFAR2 deficient cells, we repeated the same experimental set-up with infection in combination with acetate treatment on IP-10 deficient cells (IP-10-KO). Similar to FFAR2-KO cells, also IP-10-KO did not exhibit any reduction of active virus particles following acetate treatment, neither after 8 nor 24 hpi (Fig. 5G).
Taken together, acetate stimulation led to a significant reduction in viral titers at both 8 and 24 h post-infection in pulmonary epithelial cells. FFAR2 trafficking studies demonstrated that acetate promoted β-arrestin2–mediated internalization of the receptor. This antiviral effect was absent in A549 cells lacking either FFAR2 or the key interferon response mediator IP-10, suggesting both are essential for acetates mechanism during infection.
Acetate treatment affects cellular metabolism during IAV infectionBased on the in vitro results with A549 WT and FFAR2-KO cells, we concluded the beneficial effect of acetate on IAV infection is dependent on the presence of the FFAR2 receptor. Next, we aimed to further elucidate the intracellular connection between acetate and IAV infection. Since the cytokine profile from infected and stimulated human ex vivo lung slices showed no distinct alterations with acetate treatment, we proceeded to determine pro-inflammatory lipid mediators derived from arachidonic acid. Previous work has shown that cyclooxygenase products, such as prostaglandin (PG) E2, PGF2α, and thromboxane (TX) B2, are produced by macrophages during infection with IAV [30]. In the complex human ex vivo lung slice model, IAV infection resulted in an upregulation of all three mediators, most prominently for PGE2 and PGF2α. However, the acetate pre-treatment did not affect the levels of these secreted markers. Interestingly, we could not detect any lipid mediators in the supernatant of infected A549 cells (Supplemental Fig. S5B).
Furthermore, we performed mRNA transcriptomic analysis to examine the alterations caused by acetate per se in A549 cells, focusing on the transcriptomic profile of pulmonary epithelial cells with and without infection. In vitro incubation for 24 h resulted in significant changes in the overall distribution of differentially expressed genes, with a notable upregulation of 126 and downregulation of 44 genes compared to the medium control (Supplemental Fig. S5C). KEGG annotation revealed a specific upregulation of various metabolic pathways, including those involved in steroid and terpenoid biosynthesis, which are known to have antiviral functions, as determined by mRNA-sequencing (Supplemental Fig. S5D) [31].
Interestingly, at 8 h p.i., genes connected to metabolic processes such as cholesterol metabolism and glycolysis/gluconeogenesis were significantly upregulated in cells pre-treated with acetate compared to the sole infection (Supplemental Fig. S5E). Strikingly, genes involved in peroxisome function were downregulated in the same condition (Supplemental Fig. S5F). The peroxisome is known to be involved in various processes of lipid metabolism and the generation of reactive oxygen species (ROS), in addition to its involvement in viral infections [32].
In summary, acetate did not affect the secretion of lipid mediators, such as PGE2. However, 24 h pre-incubation in the pulmonary epithelial cell model resulted in an altered cell metabolism, which was significantly visible at an early point of infection. Interestingly, genes related to the peroxisome, an organelle connecting cellular metabolism and viral processes, were reduced in epithelial cells treated with acetate and infected with IAV.
Early antiviral effect of acetate during IAV infection in epithelial cellsIn the next step, we analyzed transcriptomic data from infected A549 WT cells, focusing specifically on genes related to IAV response. At 8 hpi, the genetic profile of cells infected and pre-treated with acetate (IAVAc) showed alterations compared to unstimulated infection (IAV), adjusted to the responsible mock cells (Mock and Ac). Notably, genes including IP-10 (CXCL10), Interferon induced with helicase C domain (IFIH) 1, interferon regulatory factor (IRF) 1, IRF3 or oligoadenylatsynthetase (OAS) 2 were not significantly upregulated when treated with acetate.
Further studying anti-virus-related proteins revealed that specific genes, including DDX58 and IRF9, were upregulated after 8 h. IRF9 and DDX58 are central components of the type I interferon response and play key roles in early viral sensing and antiviral signaling. Our data indicate that their expression is enhanced in acetate-treated conditions during IAV infection, suggesting that acetate may potentiate early innate immune recognition of the virus. At the later time point of 24 h, there is a general downregulation of genes related to IAV infection (Fig. 6B).
Fig. 6
Early antiviral effect of acetate during IAV infection in epithelial cells in vitro. Heatmaps of differentially expressed genes at 8 h post infection (hpi) showed more upregulated IAV response genes in acetate pre-stimulated A549 WT cells compared to controls, except for DDX58 (RIG-1) and IRF9 (A). At 24 hpi, there was again an increase in upregulated genes in infected cells pre-treated with acetate compared to controls, except for CALCOCO2, CXCL8, and JAK2 (B). Importantly, CXCL10 is the gene encoding for IP-10
When directly comparing IAVAc cells with IAV cells, various genes were significantly up- or downregulated at the early 8 h time point (Supplemental Fig. S6A). Specifically, after 8 h genes connected to toll-like receptor (TLR) and TNF signaling were significantly upregulated in IAVAc cells (Supplemental Fig. S6B). After 24 h of infection, the volcano plot—a type of scatter plot commonly used in statistics and bioinformatics to illustrate the results of differential expression or large-scale data comparisons—showed a significant upregulation and downregulation of various genes. This is represented in Supplemental Fig. S6C, which highlights the differentially expressed genes (DEGs).At this time point, the already observed reduced transcription of general IAV related was confirmed, with significantly regulated genes of the interferon and TNF signaling (Supplemental Fig. S6D).
Comments (0)